




Most studies of the effects of atmospheric CO2 enrichment on Earth's vegetation have dealt with common terrestrial plants, ranging from grasses to trees, while very few have focused on sizable plants of aquatic realms. This summary reports the results of several research studies that have evaluated the responses of marine macroalgae to elevated levels of atmospheric CO2.
In one of the earlier such studies, Gao et al. (1993) grew cultures of the red alga Gracilaria sp. and G. chilensis in vessels enriched with nitrogen and phosphorus that were continuously aerated with either normal air of 350 ppm CO2, air enriched with an extra 650 ppm CO2, or air enriched with an extra 1250 ppm CO2 for a period of 19 days. Compared to the control treatment, the relative growth enhancements in the + 650-ppm and +1250-ppm CO2 treatments were 20% and 60%, respectively, for G. chilensis, and 130% and 190%, respectively, for Gracilaria sp. Gao et al. say these results suggest that, "in their natural habitats or cultivation sites, photosynthesis and growth of Gracilaria species are likely to be CO2-limited, especially when the population density is high and water movement is slow." Thus, as the air's CO2 content continues to rise, these marine marcoalgae should be able to grow ever better; and such may well be the case with many other macroalgae too, for Gao et al. note that "photosynthesis by most macroalgae is probably limited by inorganic carbon sources in natural seawater," citing the studies of Surif and Raven (1989), Maberly (1990), Gao et al. (1991) and Levavasseur et al. (1991) as evidence for the validity of this statement.
At a glance, this latter finding might possibly be construed to imply that corals in a CO2-enriched world may be out-competed by marine macroalgae, which, to quote Langdon et al. (2003), "are not conspicuous on healthy reefs, but due to various anthropogenic pressures ... are becoming increasingly abundant." However, in an experiment they conducted at the Biosphere-2 facility near Oracle, Arizona, USA, where they studied gross primary production and calcification in a macrophyte-dominated ecosystem that had a coral cover of 3%, Langdon et al. obtained evidence that suggests that the ongoing rise in the air's CO2 content will not "hasten the transformation of reef community structure from coral to algal dominance."
Studying another red macroalga were Xu et al. (2010), who write that "Gracilaria lemaneiformis (Bory) Weber-van Bosse is an economically important red seaweed that is cultivated on a large scale in China due to the quantity and quality of agar in its cell walls." In addition, they say that "much attention has been paid to the biofiltration capacity of the species (Yang et al., 2005, 2006; Zhou et al., 2006)," and that it has thus been suggested to be "an excellent species for alleviating coastal eutrophication in China (Fei, 2004)." Considering these important characteristics of this seaweed, the authors set out to examine how this aquatic plant might respond to elevated CO2.
In conducting their experiment, plants were grown from thalli -- which were collected at 0.5 m depth from a farm located in Shen'ao Bay, Nanao Island, Shantou (China) -- for a period of 16 days in 3-L flasks of natural seawater maintained at either natural (0.5 µM) or high (30 µM) dissolved inorganic phosphorus (Pi) concentrations in contact with air of either 370 or 720 ppm CO2, while their photosynthetic rates, biomass production, and uptake of nitrate and phosphate were examined.
As best as can be determined from Xu et al.'s graphical representations of their results, algal photosynthetic rates in the natural Pi treatment were only increased by a non-significant 5% due to the 95% increase in the air's CO2 concentration, while in the high Pi treatment they were increased by approximately 41%. In the case of growth rate or biomass production, on the other hand, the elevated CO2 treatment exhibited a 48% increase in the natural Pi treatment, while in the high Pi treatment there was no CO2-induced increase in growth, due to the fact that the addition of the extra 29.5 µM Pi boosted the biomass production of the low-CO2 natural-Pi treatment by approximately 83%, and additional CO2 did not increase growth rates beyond that point.
In considering their results, the three Chinese researchers say that "elevated levels of CO2 in seawater increase the growth rate of many seaweed species despite the variety of ways in which carbon is utilized in these algae," noting that "some species, such as Porphyra yezoensis Ueda (Gao et al., 1991) and Hizikia fusiforme (Harv.) Okamura (Zou, 2005) are capable of using HCO3-, but are limited by the current ambient carbon concentration in seawater," and that "enrichment of CO2 relieves this limitation and enhances growth." With respect to the results they obtained with Gracilaria lemaneiformis, on the other hand -- which they say "efficiently uses HCO3- and whose photosynthesis is saturated at the current inorganic carbon concentration of natural seawater (Zou et al., 2004)" -- they write that "the enhancement of growth could be due to the increased nitrogen uptake rates at elevated CO2 levels," which in their experiment were 40% in the natural Pi treatment, because "high CO2 may enhance the activity of nitrate reductase (Mercado et al., 1999; Gordillo et al., 2001; Zou, 2005) and stimulate the accumulation of nitrogen, which could contribute to growth." Nevertheless, whatever strategy may be employed, these several marine macroalgae appear to be capable of benefiting greatly from increased atmospheric CO2 concentrations.
Kubler et al. (1999) grew a red seaweed common to the Northeast Atlantic intertidal zone, Lomentaria articulata, for three weeks in hydroponic cultures subjected to various atmospheric CO2 and O2 concentrations to determine the effects of these gases on growth. They report that oxygen concentrations ranging between 10 and 200% of ambient had no significant effects on daily net carbon gain or total wet biomass production rates in this seaweed. In contrast, CO2 concentrations ranging between 67 and 500% of ambient had highly significant effects these parameters. At twice the current ambient CO2 concentration, for example, daily net carbon gain and total wet biomass production rates were 52 and 314% greater than they were under ambient CO2 conditions. Even though this seaweed was likely carbon saturated, when grown at five-times the ambient CO2 concentration, it still exhibited daily net carbon gain and wet biomass production rates that were 23 and 50%, respectively, greater than those of control plants.
Zou (2005) collected specimens of a brown seaweed (Hizikia fusiforme (Harv.) Okamura) from intertidal rocks along the coast of Nanao Island, Shantou, China, and maintained them in glass aquariums in filtered natural seawater enriched with 60 µM NaNO3 and 6.0 µM NaH2PO4, where they were continuously aerated with either ambient air of 360 ppm CO2 or CO2-enriched air of 700 ppm CO2. Under these conditions he measured their relative growth and nitrogen assimilation rates, as well as their nitrate reductase activities.
The slightly less than a doubling of the air's CO2 concentration increased the seaweed's mean relative growth rate by about 50%, its mean rate of nitrate uptake during the study's 12-hour light periods by some 200%, and its nitrate reductase activity by approximately 20% over a wide range of substrate nitrate concentrations. In commenting on these findings, with respect to human health, Zou notes that "the extract of H. fusiforme has an immunomodulating activity on humans and this ability might be used for clinical application to treat several diseases such as tumors (Suetsuna, 1998; Shan et al., 1999)." He also reports that the alga "has been used as a food delicacy and an herbal ingredient in China, Japan and Korea." In fact, he says that it "is now becoming one of the most important species for seaweed mariculture in China, owing to its high commercial value and increasing market demand." As a result, the ongoing rise in the air's CO2 content bodes well for all of these applications. In addition, Zou notes that "the intensive cultivation of H. fusiforme would remove nutrients more efficiently with the future elevation of CO2 levels in seawater, which could be a possible solution to the problem of ongoing coastal eutrophication," suggesting that rising CO2 levels may also assist in the amelioration of this environmental problem.
Palacios and Zimmerman (2007) conducted an experiment on eelgrass (Zostera marina) at the Duke Energy-North America Power Plant at Moss Landing, California (USA), where flue gas generated by the power plant furnace was piped approximately 1 km to a site where it was bubbled through outdoor flow-through seawater aquaria at rates that produced four different aqueous CO2 treatments characteristic of: "(1) the present day atmosphere, with approximately 16 µM CO2(aq), (2) CO2 projected for 2100 that increases the CO2(aq) concentration of seawater to approximately 36 µM CO2(aq), (3) CO2 projected for 2200 that increases the CO2(aq) concentration of seawater to 85 µM CO2(aq), and (4) a dissolved aqueous CO2 concentration of 1123 µM CO2(aq), which triples the light-saturated photosynthesis rate of eelgrass (Zimmerman et al., 1997)."
Results indicated that the elevated CO2 "led to significantly higher reproductive output, below-ground biomass and vegetative proliferation of new shoots in light-replete treatments," i.e., those receiving light at 33% of the surface irradiance level. More specifically, they write that "shoots growing at 36 µM CO2(aq) were 25% larger than those in the unenriched treatment [16 µM CO2(aq)]," while "at 85 µM CO2(aq) shoots were 50% larger than those in the unenriched treatment and at 1123 µM CO2(aq) shoots were almost twice as large as those in the unenriched treatment." In addition, they found that at 1123 µM CO2(aq) "22% of the shoots differentiated into flowers, more than twice the flowering output of the other treatments at this light level."
These findings are of great significance and have far-reaching implications. Noting that "increased CO2(aq) is capable of increasing eelgrass reproductive output via flowering, and area-specific productivity via vegetative shoot proliferation under naturally replete light regimes," Palacios and Zimmerman state that "the resulting increases in eelgrass meadow density may initiate a positive feedback loop that facilitates the trapping of sediments and prevents their resuspension, thereby reducing turbidity and increasing light penetration in coastal habitats," such that the resulting increased light penetration "may allow seagrass colonization depths to increase even further."
The two researchers also suggest that the CO2-induced increase in the productivity of eelgrass may "enhance fish and invertebrate stocks as well." In fact, they go so far as to suggest that the "deliberate injection of CO2 to seawater may facilitate restoration efforts by improving the survival rates of recently transplanted eelgrass shoots," noting that "it can buffer the negative effects of transplant shock by increasing rhizome reserve capacity and promoting shoot proliferation in light-replete environments." In addition, they say it "may also facilitate eelgrass survival in environments where conditions are periodically limiting, such as long dark winters or unusually warm summers that produce unfavorable productivity to respiration ratios," and they state that "CO2 injection may also promote flowering and seed production necessary for expansion and maintenance of healthy eelgrass meadows." What is more, they suggest that "rising concentrations of CO2(aq) may increase vegetative propagation and seed production of other seagrass populations besides eelgrass."
According to Suarez-Alvarez et al. (2012), "the increase of coastal activities which produce important amounts of wastes, including inorganic nutrients (Troell et al., 2003), makes micro- and macroalgae interesting organisms to predict possible impacts, responses, and remediation processes by considering biomass production through cultivation techniques," citing Gao et al. (1991, 1993), Gao and McKinley (1994), Keffer and Kleinheinz (2002), Doucha et al. (2005) and Israel et al. (2005). And they say that "macroalgae, in particular, have been the object of additional interest for CO2 remediation (Gao and McKinley, 1994) because of their solar energy conversion capacity, high productivity values (higher than most productive terrestrial crops) and the possibility of being, in many cases, intensively cultivated."
In a further assessment these possibilities and how they might be impacted by atmospheric CO2 enrichment (750 and 1600 ppm compared to an ambient value of 360 ppm), Suarez-Alvarez et al. cultivated 8-gram fragments of the macroalga Hypnea spinella in 1-L flasks containing filtered seawater enriched with 140 µM NH4Cl and 14 µM KH2PO4 for 7 days of acclimation, after which they were culled to their initial density and grown for 9 more days, during which period the three researchers measured various plant physiological properties and processes.
In the case of light-saturated net photosynthesis, the authors report that rates in the 750-ppm and 1600-ppm CO2 treatments were 41.5% and 50.5% greater, respectively, than what was measured in the 360-ppm treatment, while relative growth enhancements were 85.6% and 63.2% greater, respectively, and maximum ammonium uptake rates were enhanced by 24.2% and 19.9%, respectively, in the 750- and 1600-ppm CO2 treatments. "From a practical point of view," in the words of Suarez-Alvarez et al., "these results suggest that intensive culture of H. spinella operated in biofilters might be enhanced by CO2 supply to generate higher biomass productivities and better nitrogen biofiltration efficiencies," and they say that "the use of flue gases for this purpose would also improve the ability of bioremediation of these biofilters, as has already been tested for Gracilaria cornea," citing Israel et al. (2005). And thus it is that what some have called an air pollutant (CO2) may ironically soon play the role of a cleanser of water pollutants.
Writing as background for their study, Jiang et al. (2010) note that "seagrasses are flowering plants that thrive in shallow oceanic and estuarine waters around the world, and are ranked as one of the most ecologically and economically valuable biological systems on Earth," citing the work of Beer et al. (2006). And they say that Thalassia hemprichii "is among the most widely-distributed seagrass species in an Indo-Pacific flora, dominating in many mixed meadows," citing the work of Short et al. (2007). Against this backdrop, Jiang et al. collected intact vegetative plants of T. hemprichii from Xincun Bay of Hainan Island, Southern China, which they transported to the laboratory and cultured in flow-through seawater aquaria bubbled with four different concentrations of CO2 that were representative of (1) the present global ocean, with a pH of 8.10, (2) the projected ocean for 2100, with a pH of 7.75, (3) the projected ocean for 2200, with a pH of 7.50, and (4) the ocean characteristic of "an extreme beyond the current predictions" (a 100-fold increase in free CO2, with a pH of 6.2).
With respect to the results of their analysis, the three researchers report that the "leaf growth rate of CO2-enriched plants was significantly higher than that in the unenriched treatment," that "nonstructural carbohydrates (NSC) of T. hemprichii, especially in belowground tissues, increased strongly with elevated CO2," and that "belowground tissues showed a similar response with NSC."
As for the implications of their findings, i.e., that "CO2 enrichment enhances photosynthetic rate, growth rate and NSC concentrations of T. hemprichii," the Chinese scientists list several. They say that with higher atmospheric CO2 concentrations "colonization beyond current seagrass depth limits is possible," that the extra stored NSC "can be used to meet the carbon demands of plants during periods of low photosynthetic carbon fixation caused by severe environmental disturbance such as underwater light reduction," that it can enhance "rhizome growth, flowering shoot production and vegetative proliferation," and that it "may buffer the negative effects of transplant shock by increasing rhizome reserve capacity." They also write that "the globally increasing CO2 may enhance seagrass survival in eutrophic coastal waters, where populations have been devastated by algal proliferation and reduced column light transparency," and that "ocean acidification will stimulate seagrass biomass and productivity, leading to more favorable habitat and conditions for associated invertebrate and fish species."
In a review paper, Wu et al. (2008) listed several known effects of a CO2-enriched atmosphere on micro- and macro-algae living in the world's oceans. According to the authors, "enriched CO2 up to several times the present atmospheric level has been shown to enhance photosynthesis and growth of both phytoplanktonic and macro-species that have less capacity of CCMs [CO2-concentrating mechanisms]," adding that "even for species that operate active CCMs and those whose photosynthesis is not limited by CO2 in seawater, increased CO2 levels can down-regulate their CCMs and therefore enhance their growth under light-limiting conditions," because "at higher CO2 levels, less light energy is required to drive CCM." In addition, they report that enhanced CO2 levels have been found to enhance the activity of nitrogen reductase in several marine plants, and that this phenomenon "would support enhanced growth rate by providing adequate nitrogen required for the metabolism under the high CO2 level." Last of all, they say that "altered physiological performances under high-CO2 conditions may cause genetic alteration in view of adaptation over long time scales," and that "marine algae may adapt to a high CO2 oceanic environment so that evolved communities in [the] future are likely to be genetically different from contemporary communities."
In a somewhat different type of study, Semesi et al. (2009) investigated whether diel variations in seawater pH caused by the photosynthetic activity of seagrass meadows within Chwaka Bay (Zanzibar, Tanzania) could affect the calcification and photosynthesis rates of calcareous red algae (Hydrolithon sp. and Mesophyllum sp.) and green algae (Halimeda renschii) growing within the marine meadows. This feat was accomplished by measuring rates of calcification and relative photosynthetic electron transport (rETR) of the algae in situ in open-bottom incubation cylinders either in the natural presence of the rooted seagrasses or after their leaves had been removed.
Based on their investigation, Semesi et al. report that "seagrass photosynthesis increased the seawater pH within the cylinders from 8.3-8.4 to 8.6-8.9 after 2.5 hours (largely in conformity with that of the surrounding seawater), which, in turn, enhanced the rates of calcification 5.8-fold for Hydrolithon sp. and 1.6-fold for the other 2 species," while "the rETRs of all algae largely followed the irradiance throughout the day and were (in Mesophyllum sp.) significantly higher in the presence of seagrassses." The three researchers therefore conclude that "algal calcification within seagrass meadows such as those of Chwaka Bay is considerably enhanced by the photosynthetic activity of the seagrasses, which in turn increases the seawater pH." In fact, they state that the high pH values that were created by seagrass photosynthesis are what actually caused the elevated calcification rates of the macroalgae; and this observation has some further implications. It suggests, for example, that the lowering of seawater pH caused by higher atmospheric CO2 concentrations -- which results in a lowering of the calcium carbonate saturation state of seawater, and which has been predicted to lead to reductions in coral calcification rates -- may well be counteracted by CO2-induced increases in the photosynthetic activity of the symbiotic zooxanthellae of Earth's corals, which phenomenon may boost the pH of seawater in intimate contact with the coral host, allowing the host to actually increase its calcification rate in CO2-enriched seawater (or to at least not suffer a major reduction in calcification).
Hurd et al. (2011) observe that "most ocean acidification studies so far have been simplistic," in that they have not "jointly considered physical, chemical and biological interactions." They note, however, that "the emerging discipline of marine ecomechanics (Denny and Helmuth, 2009; Denny and Gaylord, 2010) provides a valuable framework in which such inter-disciplinary research can be conducted." The old experimental approach, as they describe it, "overlooks the existence of a discrete micro-layer (i.e., diffusion boundary layer, DBL) at the surface of many aquatic organisms that buffers them from the surrounding mainstream seawater (Vogel, 1996)." This feat is achieved by metabolic processes that alter the water chemistry within the DBL, with photosynthesis increasing pH, and calcification and respiration reducing pH (Hurd et al., 2009). Therefore, as they continue, "the chemical environment within the DBL differs from that in the mainstream seawater just micrometers away, with implications for both the dissolution of, and formation of, calcium carbonate (Borowitzka and Larkum, 1976; Ries et al., 2009)."
In a study employing the still-evolving ecomechanic approach, Hurd et al. used pH micro-electrodes and oxygen micro-optodes to directly measure the DBL thickness at the surface of the coralline seaweed Sporolithon durum at a range of seawater velocities (0-10 cm/sec) that reflected those found within a temperate reef in Southern New Zealand (45.38°S) that may be vulnerable to ocean acidification (OA). They also determined whether or not DBL thickness would be affected when mainstream seawater pH was reduced to 7.5, which is the projected worst-case scenario for the year 2215 as calculated by Caldeira and Wickett (2003). In addition, they measured pH fluctuations at the surface of S. durum on a timescale of hours at ambient seawater pH and pH 7.5 at two different flows (1.5 and 6.3 cm/sec).
The seven scientists determined that coralline seaweeds encounter a wide range of pH values over each daily cycle; but they found that they are able to increase their pH substantially due to photosynthesis and to successfully withstand periods of very low pH (relative to the present day and comparable to values predicted for coming centuries) under low flows. In concluding their report, Hurd et al. say their findings "support the view that although the role of chemistry on OA is well understood, the biological responses to OA will be complex," citing their own work and that of Fabry et al. (2008), while noting that "both the site of calcification and the ecomechanics of the biota, i.e., the interactions between their morphology, physiology and the surrounding hydrodynamic environment, must be considered."
Investigating the same topic, but three years later, Cornwall et al. (2014) state that "recent studies suggest that OA could be less detrimental to calcifying species that photosynthesize, such as coralline algae, growing in slow-flow compared to fast-flow habitats ... because the diffusion boundary layer (hereafter 'DBL') provides a thin but significant biologically-controlled buffer between the calcifying organism's external structure and the outer bulk seawater, potentially reducing rates of dissolution (Hurd et al., 2011; Cornwall et al., 2013.)." To further explore this possibility, Cornwall et al. grew the coralline macroalga Arthrocardia corymbosa in a multi-factorial experiment with two mean pH levels (8.05 'ambient' and 7.65 a worst case 'ocean acidification' scenario projected for 2100), each with two rates of seawater flow: either fast or slow, producing either thin or thick DBLs, respectively.
The eight researchers report that "growth under ambient seawater pH (8.05) was not significantly different in thin and thick DBL treatments," but they found that "coralline algae grown under slow flows with thick DBLs maintained net growth and calcification at pH 7.65, whereas those in higher flows with thin DBLs had net dissolution." And, in light of what they learned, Cornwall et al. conclude that low-flow conditions that promote the formation of thick DBLs may enhance the subsistence of various calcifiers by creating localized hydrodynamic conditions "where metabolic activity ameliorates the negative impacts of ocean acidification." And they thus propose the possibility of "maintaining seaweed beds characterized by localized biological modification of pH and seawater flow, as potential refugia from OA."
Anthony et al. (2011) used "a carbon flux model for photosynthesis, respiration, calcification and dissolution coupled with Lagrangian transport to examine how key groups of calcifiers (zooxanthellate corals) and primary producers (macroalgae) on coral reefs contribute to changes in the seawater carbonate system as a function of water residence time." This work revealed, in their words, that "the carbon fluxes of corals and macroalgae drive Ωa in opposing directions," such that "areas dominated by corals elevate pCO2 and reduce Ωa, thereby compounding ocean acidification effects in downstream habitats, whereas algal beds draw CO2 down and elevate Ωa, potentially offsetting ocean acidification impacts at the local scale." And they also report that simulations for two significantly elevated CO2 scenarios (600 and 900 ppm CO2) suggested that "a shift in reef community composition from coral to algal dominance in upstream areas under ocean acidification will potentially improve conditions for calcification in downstream areas."
Field validation of the simulations of Anthony et al. was provided by Kleypas et al. (2011), who examined the roles of three key members of benthic reef communities (corals, macroalgae and sand) in modifying the chemistry of open-ocean source water, finding that "the drawdown of total dissolved inorganic carbon due to photosynthesis and calcification of reef communities can exceed the drawdown of total alkalinity due to calcification of corals and calcifying algae, leading to a net increase in aragonite saturation state." In addition, they note that there were no seagrasses on the reef flat they studied; and they state that "research suggests that seagrasses may have an additional impact on reef seawater chemistry because they enhance the alkalinity flux from sediments (Burdige and Zimmerman, 2002), and they respond to CO2 fertilization (Palacios and Zimmerman, 2007)."
In light of these several observations, it might logically be expected that reef communities would gradually alter their spatial compositions in a CO2-acreting world to the point where seagrasses and other macroalgae take up residence in upstream regions, while corals and other calcifying organisms lay claim to downstream regions. Therefore, as Anthony et al. conclude, "although the carbon fluxes of benthic reef communities cannot significantly counter changes in carbon chemistry at the scale of oceans, they provide a significant mechanism of buffering ocean acidification impacts at the scale of habitat to reef."
In a discussion of the same topic, Manzello et al. (2012) write that although many people expect future ocean acidification (OA) due to rising atmospheric CO2 concentrations to reduce the calcification rates of marine organisms, we have little understanding of how OA will manifest itself within dynamic, real-world systems, because, as they correctly note, "natural CO2, alkalinity, and salinity gradients can significantly alter local carbonate chemistry, and thereby create a range of susceptibility for different ecosystems to OA." Against this backdrop, in an effort "to determine if photosynthetic CO2 uptake associated with seagrass beds has the potential to create OA refugia," as they describe it, Manzello et al. repeatedly measured carbonate chemistry across an inshore-to-offshore gradient in the upper, middle and lower Florida Reef Tract over a two-year period.
During times of heightened oceanic vegetative productivity, the five U.S. researchers found "there is a net uptake of total CO2 which increases aragonite saturation state (Ωarag) values on inshore patch reefs of the upper Florida Reef Tract," and they say that "these waters can exhibit greater Ωarag than what has been modeled for the tropical surface ocean during preindustrial times, with mean Ωarag values in spring equaling 4.69 ± 0.10." At the same time, however, they report that ?arag values on offshore reefs "generally represent oceanic carbonate chemistries consistent with present day tropical surface ocean conditions."
Manzello et al. hypothesize that the pattern described above "is caused by the photosynthetic uptake of total CO2 mainly by seagrasses and, to a lesser extent, macroalgae in the inshore waters of the Florida Reef Tract." And they therefore conclude that these inshore reef habitats are "potential acidification refugia that are defined not only in a spatial sense, but also in time, coinciding with seasonal productivity dynamics," which further implies that "coral reefs located within or immediately downstream of seagrass beds may find refuge from ocean acidification." And in further support of this conclusion, they cite the work of Palacios and Zimmerman (2007), which they describe as indicating that "seagrasses exposed to high-CO2 conditions for one year had increased reproduction, rhizome biomass, and vegetative growth of new shoots, which could represent a potential positive feedback to their ability to serve as ocean acidification refugia."
Introducing their study, Johnson et al. (2012) write that "CO2 gradients in natural settings, where whole ecosystems have been exposed to elevated levels of CO2, allow us to investigate changes in the interactions, competition, predation and/or herbivory that involve long-lived metazoan species in benthic marine ecosystems." And they note, in this regard, that "volcanic CO2 gradients are beginning to reveal the ecological shifts that can be expected to occur with globally increasing atmospheric CO2 in both temperate (Hall-Spencer et al., 2008) and tropical ecosystems (Fabricius et al., 2011)."
Against this backdrop Johnson et al. assessed the abundance of herbivores (sea urchins) and the response of brown macroalgae (Padina spp.) to increasing levels of CO2 in two natural settings. One of the sites of their research was a set of shallow, volcanic CO2 seeps on the island of Vulcano, NE Sicily, where P. pavonica was studied; while the other site, where P. australis was studied, took place at comparable seeps in the D'Entrecasteaux Island group, Papua New Guinea.
The five scientists report that "there was a reduction in sea urchin abundances alongside a proliferation of Padina spp., as CO2 levels increased" along both temperate and tropical rocky shores. In the case of sea urchins, in fact, they discovered that the predators were actually absent in locations having the highest CO2 levels (lowest pH); while in the case of the Padina spp., they found that "even in the lowest pH conditions, P. pavonica and P. australis were still able to calcify, seemingly from the enhancement of photosynthesis under high levels of CO2."
In considering their findings, Johnson et al. opine that the absence of sea urchins in the CO2-enriched areas "may be one explanation for the proliferation of Padina spp., as it becomes released from the top-down control by these keystone grazers," noting that "this effect of sea urchin removal has been observed in other Padina sp. populations (Sammarco et al., 1974) and across other Phaeophyte assemblages (Leinaas and Christie, 1996; Ling et al.,2010)," while in regard to the increase they observed in the photosynthetic capacity of the Padina species under conditions of higher CO2, they note that "increased photosynthetic activity at high CO2 has also been observed in other calcified macroalgae (Reiskind et al., 1988; Semesi et al., 2009)," as well as in "non-calcified macroalgae (Kubler et al., 1999; Connell and Russell, 2010; Russell et al., 2011)."
Egilsdottir et al. (2013) present the rationale for their work by writing "it has been suggested that organisms presently surviving in highly variable environments are likely to be more robust to ocean acidification," citing Moulin et al. (2011) and Raven (2011); and they suggest that if this be the case, "the ability of organisms to tolerate significant pH/pCO2 fluctuations may be a result of adaptation (a genetic trait shared by the population) and/or acclimation (owing to phenotypic plasticity of the individual)."
"To test this hypothesis," in the words of Egilsdottir et al., "the effect of elevated pCO2 was investigated in the articulated coralline red alga Corallina elongata from an intertidal rock pool on the north coast of Brittany (France), where pCO2 naturally varied daily between 70 and 1000 µatm." This was done by growing the algae at four different pCO2 values (380, 550, 750 and 1000 µatm) in laboratory mesocosms and measuring several different physiological responses of the coral.
In discussing their results, the French and Icelandic researchers report that "algae grown under elevated pCO2 formed fewer new structures and produced calcite with a lower mMg/Ca ratio relative to those grown under 380 µatm." However, they also report that "respiration, gross primary production and calcification in light and dark were not significantly affected by increased pCO2." Egilsdottir et al. conclude that their study "supports the assumption that C. elongata from a tidal pool, where pCO2 fluctuates over diel and seasonal cycles, [are] relatively robust to elevated pCO2." And they also thus state that these observations portend "a greater resilience" of such organisms to the projected "highly variable pH/pCO2 environments of future ocean acidification."
In a study designed to determine if a set of species-specific "tipping points" might exist in the calcification responses of coral reef calcifiers to increasing ocean acidification (OA) driven by elevated atmospheric CO2 concentrations, Comeau et al. (2013) "compared the effects of six partial pressures of CO2 from 28 Pa to 210 Pa on the net calcification of four corals (Acropora pulchra, Porites rus, Pocillopora damicornis, and Pavona cactus), and four calcified algae (Hydrolithon onkodes, Lithophyllum flavescens, Halimeda macroloba, and Halimeda minima)." This they did from August to October of 2011 in Moorea, French Polynesia, using organisms collected from the back reef of the island's north shore at 1-2 meters depth, which they placed in "a mesocosm apparatus consisting of 12 tanks that allowed for the maintenance of six PCO2 treatments in duplicate," where the calcifiers were studied in detail for several 2-week periods.
Results indicated that calcification rates in two of the eight species (P. damicornis and H. macroloba) were impervious to changes in pH across the full range of treatment. As for the remaining six species, they experienced a mean decrease in net calcification of ~10% when the ambient PCO2 (39 Pa) was doubled. However, all of these species maintained positive rates of calcification up through the highest PCO2, where the calcium carbonate aragonite saturation sate was approximately equal to 1.
In discussing the significance of their findings, Comeau et al. write that "in contrast to previous studies that have predicted rapid decreases in calcification of corals and coral reefs exposed to PCO2 ≥ 50 Pa ... our study, performed at the organismic level on eight of the main calcifiers in Moorea, suggests that tropical reefs might not be affected by OA as strongly or as rapidly as previously supposed" ... or not at all in the case of some species, it might be added, in light of the nil responses of P. damicornis and H. maeroloba noted above. Further, when PCO2 was increased to 210 Pa (fully double the pessimistic value predicted by some for the end of the century), they say they "did not detect a threshold at which the effect of PCO2 on calcification became nonlinear and intensified (i.e., a tipping point)."
Moving closer to the present, Comeau et al. (2014a) conducted a study of two coral taxa and two calcifying algae, which they conducted in Moorea (French Polynesia), Hawaii (USA) and Okinawa (Japan). Their work revealed that for three of the four calcifiers "there was no effect of pCO2 on net calcification" at any of the three locations, which led them to suggest that this finding "may represent a constitutive and geographically conserved capacity to resist some of the effects of OA."
Introducing their work, Garrard and Beaumont (2014) state that seagrass beds provide numerous important ecosystem services, such as "protection of the coastline, bioremediation of waste, food provision and maintenance of marine biodiversity," citing the works of Jackson et al. (2012) and Cullen-Unsworth and Unsworth (2013). And in light of these facts, they review how the several mentioned services are likely to be impacted by the ongoing rise in the atmosphere's CO2 concentration.
Based on projections of future anthropogenic CO2 emissions and their impacts on the above- and below-ground growth of seagrass, the two UK researchers estimate that over the remainder of this century, the global standing stock of seagrass "is expected to increase by 94%, whilst the standing stock in the UK is expected to increase by 82%." And they calculate that the associated value of this increase in both above- and below-ground carbon sequestration capacity is -- when summed over the entire world -- approximately 500 and 600 billion pounds sterling ($765-918 billion USD), respectively, between 2010 and 2100. They thus conclude that "sustainable management of seagrasses is critical to avoid their continued degradation and loss of carbon sequestration capacity," and, it might be added, to maintain -- or even enhance -- their many important ecosystem services.
Noting that organisms living near natural volcanic seeps are often acclimatized -- and in some cases even adapted -- to elevated CO2 environments, Vogel et al. (2015) write that volcanic CO2 seeps thus provide an opportunity to identify which organisms are capable of living in elevated CO2 conditions projected to occur globally just a few decades from now. And, therefore, they investigated various ecological, physiological and skeletal characteristics of the calcifying green alga Halimeda growing in the vicinity of natural CO2 seeps (pH ~7.8) and compared them to those observed at control reefs exhibiting ambient CO2 conditions (pH ~8.1).
This work revealed, as the six scientists report, that for each of the two most abundant species (H. digitata and H. opuntia), rates of calcification in the light increased at the high CO2 site compared to controls by 131% and 41%, respectively. In the dark, on the other hand, they say that "calcification was not affected by elevated CO2 in H. digitata, whereas it was reduced by 167% in H. opuntia, suggesting nocturnal decalcification." However, they also found that 24-hour net calcification was similar between seep and control sites, implying that the increased calcification when exposed to normal daylight more than compensated for the reduced dark calcification in H. opuntia.
Based on the findings of their study of "long-term exposure in a natural environment with natural light, nutrient and flow regimes," Vogel et al. thus conclude -- in the final sentence of their paper's abstract -- that the Halimeda spp. they investigated "can acclimatize and will likely still be capable to grow and calcify in PCO2 conditions exceeding most pessimistic future CO2 projections."
Introducing their publication in Marine Biology, Ow et al. (2015) note that increased atmospheric carbon dioxide leads to ocean acidification (OA) and CO2 enrichment of seawater, which leads them to further remark that "given the important ecological functions of seagrass meadows, understanding their responses to CO2 will be critical for the management of coastal ecosystems."
Following the implications of their own assessment of this still-evolving situation, the three Australian researchers describe how they examined the physiological responses of three tropical seagrasses (Cymodocea serrulata, Halodule uninervis and Thalassia hemprichii) to a range of seawater pCO2 levels in a laboratory study, where plants of the three species "were exposed to four different pCO2 treatments (442-1204 µatm) for two weeks, approximating the range of end-of-century emission scenarios." And what did they thereby learn?
Ow et al. say the results of their study "demonstrated that tropical seagrasses can increase their photosynthetic rates, adjust photosynthetic performance and increase growth rates in response to CO2 enrichment" (Figure 1), and that this "ability of marine macro-autotrophs to utilize the greater CO2 availability suggests that they will thrive under future scenarios of climate change," further citing Koch et al. (2013) in this regard and adding that still-earlier "observations of high seagrass abundance at CO2 seep sites indicate seagrass productivity might continually benefit from pCO2 enrichment over the long term," citing Fabricius et al. (2011).
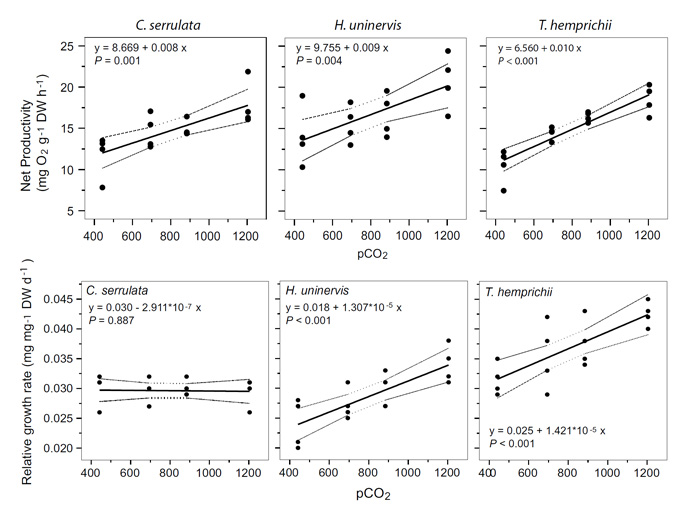
Figure 1. Linear model fits (dotted lines indicate 95 % confidence intervals) for net productivity and relative growth rates of C. serrulata, H. uninervis and T. hemprichii in response to pCO2 enrichment.
Introducing the subject of their study, Pajusalu et al. (2015) write that charophytes are "submerged, rooted algae with well-developed complex thalli and morphology" that grow in both marine and fresh waters, additionally noting that "charophyte communities are important habitats for a number of invertebrate species and epiphytic algae and they provide feeding and nursery areas for several species of fish and birds," citing the works of Martin et al. (2003) and Torn (2008). Yet despite these important functions, the authors say the response of charophyte communities to increasing CO2 concentrations, i.e., ocean acidification, "is not known."
In an effort to remedy this situation, Pajusalu et al. conducted an experiment to measure the net primary production of three charophyte species (Chara aspera, Chara horrida and Chara tomentosa) exposed to three different pCO2 levels (approximately 2000, 1000 and 200 µatm) over a period of eight days. The experiment was performed during the summer in field mesocosms constructed in a shallow bay located in the Gulf of Riga, northeast Baltic Sea (58.37°N, 22.98°E).
As illustrated in Figure 2, net primary production (NPP) varied between charophyte species and pCO2 treatment level, where higher pCO2 stimulated NPP from 5 to 50 percent above ambient (~200 µatm and described by the authors as "natural" for this region of the sea). And as a result of these findings, they note in the conclusion of their paper that "increased CO2 levels in brackish water may enhance the photosynthetic activity of charophytes in shallow coastal ecosystems under summer conditions," since "the natural content of CO2 in brackish water most likely limits the primary production of charophytes."
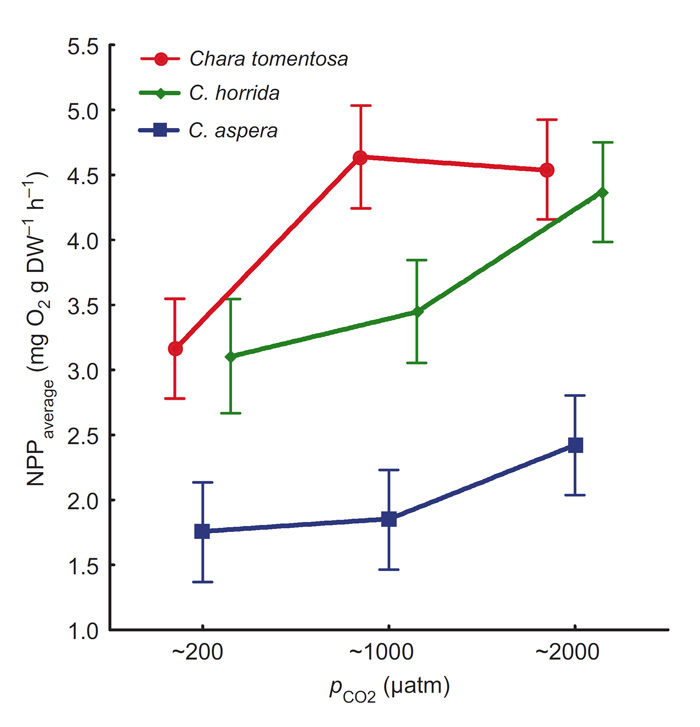
Figure 2. Mean net primary production (NPP) rates (n = 24, error bars are ± 95% CI) measured during the entire eight-day experimental period for the three tested charophyte species at different brackish-water pCO2 levels. Adapted from Pajusalu et al. (2015).
In one final study, noting that "predicted changes in seawater carbonate chemistry can influence important biological and physiological processes of calcifying and non-calcifying marine organisms" and that "seaweeds are the base of the food web and major contributors to benthic primary production in coastal ecosystems," Fernández et al. (2015) designed an experiment to study the effects of declining oceanic pH on Macrocystis pyrifera, a "dominant, highly productive brown seaweed of temperate coastlines of the northern and southern Pacific."
To accomplish their objective, samples of Macrocystis were acquired from Otago Harbor, New Zealand and then grown in a laboratory for seven days under present day and predicted future atmospheric CO2 concentrations, which corresponded to seawater pH treatments of 8.05 and 7.59, respectively. Among their reported findings, the three researchers note there was no significant difference in photosynthetic rates of Macrocystis growing at either pH treatment level. Similarly, there was no significant difference in relative growth rate, although growth rates measured on days 3 and 7 averaged 28% and 17% higher, respectively, in the future pH treatment (pH of 7.59) than the present-day treatment (pH of 8.05) (Figure 3).
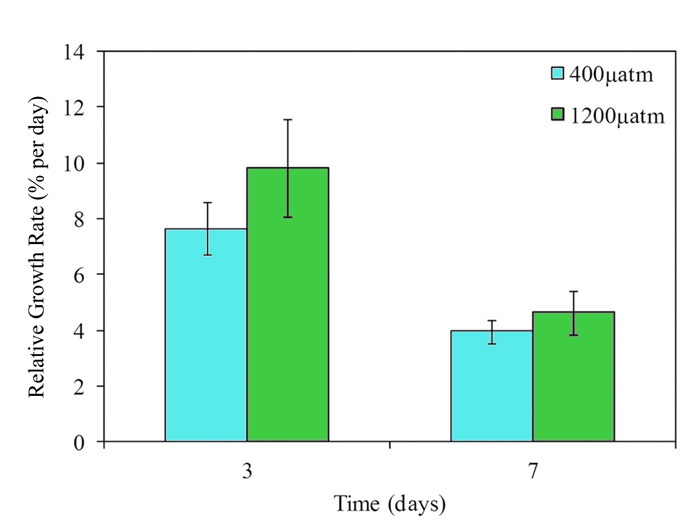
Figure 3. Relative growth rate of Macrocystis pyrifera measured on days 3 and 7 of pH treatment.
The results of Fernández et al.'s work -- plus the many other studies cited in this report above -- are encouraging in that they contrast with the seemingly never ending barrage of news stories predicting gloom and doom for marine life from ocean acidification. As demonstrated here, for many species, ocean acidification will be a non-problem ... and maybe even a blessing!
References
Anthony, K.R.N., Kleypas, J.A. and Gattuso, J.-P. 2011. Coral reefs modify their seawater carbon chemistry -- implications for impacts of ocean acidification. Global Change Biology 10.1111/j.1365-2486.2011.02510.x.
Beer, S., Mtolera, M., Lyimo, T. and Bjork, M. 2006. The photosynthetic performance of the tropical seagrass Halophila ovalis in the upper intertidal. Aquatic Botany 84: 367-371.
Borowitzka, M.A. and Larkum, A.W.D. 1976. Calcification in the green alga Halimeda. Journal of Experimental Botany 27: 879-893.
Comeau, S., Carpenter, R.C., Njiri, Y., Putnam, H.M., Sakai, K. and Edmunds, P.J. 2014a. Pacific-wide contrast highlights resistance of reef calcifiers to ocean acidification. Proceedings of the Royal Society B 281: 10.1098/rspb.2014.1339.
Comeau, S., Edmunds, P.J., Spindel, N.B. and Carpenter, R.C. 2013. The responses of eight coral reef calcifiers to increasing partial pressure of CO2 do not exhibit a tipping point. Limnology and Oceanography 58: 388-398.
Connell, S.D. and Russell, B.D. 2010. The direct effects of increasing CO2 and temperature on non-calcifying organisms: increasing the potential for phase shifts in kelp forests. Proceedings of the Royal Society of London B 277: 1409-1415.
Cornwall, C.E., Boyd, P.W., McGraw, C.M., Hepburn, C.D., Pilditch, C.A., Morris, J.N., Smith, A.M. and Hurd, C.L. 2014. Diffusion boundary layers ameliorate the negative effects of ocean acidification on the temperate coralline macroalga Arthrocardia corymbosa. PLOS ONE 9: e97235.
Cornwall, C.E., Hepburn, C.D., Pilditch, C.A. and Hurd, C.L. 2013. Concentration boundary layers around complex assemblages of macroalgae: Implications for the effects of ocean acidification on understory coralline algae. Limnology and Oceanography 58: 121-130.
Cullen-Unsworth, L and Unsworth, R. 2013. Seagrass meadows, ecosystem services, and sustainability. Environment 55: 14-27.
Denny, M.W. and Gaylord, B. 2010. Marine ecomechanics. Annual Review of Marine Science 2: 89-114.
Denny, M. and Helmuth, B. 2009. Grand challenges. Confronting the physiological bottleneck: a challenge from ecomechanics. Integrated Comparative Biology 49: 197-201.
Doucha, J., Straka, F. and Livansky, K. 2005. Utilization of flue gas for cultivation of microalgae (Chlorella sp.) in an outdoor open thin-layer photobioreactor. Journal of Applied Phycology 17: 403-412.
Egilsdottir, H., Noisette, F., Noel, L.M.-L.J., Olafson, J. and Martin, S. 2013. Effects of pCO2 on physiology and skeletal mineralogy in a tidal pool coralline alga Corallina elongata. Marine Biology 160: 2103-2112.
Fabricius, K.E., Langdon, C., Uthicke, S., Humphrey, C., Noonan, S., De'ath, G., Okazaki, R., Koch, M, Bowes, G., Ross, C. and Zhang, X.-H. 2013. Climate change and ocean acidification effects on seagrasses and marine macroalgae. Global Change Biology 19: 103-132.
Fabricius, K.E., Langdon, C., Uthicke, S., Humphrey, C., Noonan, S., De'ath, G., Okazaki, R., Muehllehner, N., Glas, M.S. and Lough, J.M. 2011. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nature Climate Change 1: 165-169.
Fabry, V.J., Seibel, B.A., Feely, R.A. and Orr, J.C. 2008. Impacts of ocean acidification on marine fauna and ecosystem processes. ICES Journal of Marine Science 65: 414-432.
Fei, X.G. 2004. Solving the coastal eutrophication problem by large scale seaweed cultivation. Hydrobiologia 512: 145-151.
Fernández, P.A., Roleda, M.Y. and Hurd, C.L. 2015. Effects of ocean acidification on the photosynthetic performance, carbonic anhydrase activity and growth of the giant kelp Macrocystis pyrifera. Photosynthesis Research 124: 293-304.
Gao, K., Aruga, Y., Asada, K., Ishihara, T., Akano, T. and Kiyohara, M. 1991. Enhanced growth of the red alga Porphyra yezoensis Ueda in high CO2 concentrations. Journal of Applied Phycology 3: 355-362.
Gao, K., Aruga, Y., Asada, K. and Kiyohara, M. 1993. Influence of enhanced CO2 on growth and photosynthesis of the red algae Gracilaria sp. and Gracilaria chilensis. Journal of Applied Phycology 5: 563-571.
Gao, K. and McKinleuy, K.R. 1994. Use of macroalgae for marine biomass production and CO2 remediation: a review. Journal of Applied Phycology 6: 45-60.
Garrard, S.L. and Beaumont, N.J. 2014. The effect of ocean acidification on carbon storage and sequestration in seagrass beds: a global and UK context. Marine Pollution Bulletin 86: 138-146.
Gordillo, F.J.L., Niell, F.X. and Figueroa, F.L. 2001. Non-photosynthetic enhancement of growth by high CO2 level in the nitrophilic seaweed Ulva rigida C. Agardh (Chlorophyta). Planta 213: 64-70.
Hall-Spencer, J.M., Rodolfo-Metalpa, R., Martin, S., Ransome, E., Fine, M., Turner, S.M., Rowley, S.J., Tedesco, D. and Buia, M.-C. 2008. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 454: 96-99.
Hurd, C.L., Cornwall, C.E., Currie, K., Hepburn, C.D., McGraw, C.M., Hunter, K.A. and Boyd, P.W. 2011. Metabolically induced pH fluctuations by some coastal calcifiers exceed projected 22nd century ocean acidification: a mechanism for differential susceptibility? Global Change Biology 17: 3254-3262.
Hurd, C.L., Hepburn, C.D., Currie, K.I., Raven, J.A. and Hunter, K.A. 2009. Testing the effects of ocean acidification on algal metabolism: considerations for experimental designs. Journal of Phycology 45: 1236-1251.
Israel, A.,Gavrieli, J., Glazer, A. and Friedlander, M. 2005. Utilization of flue gas from a power plant for tank cultivation of the red seaweed Gracilaria cornea. Aqaculture 249: 311-316.
Jackson, E.L., Langmead, O., Beaumont, N., Potts, T. and Hattam, C.A. 2012. Seagrass Ecosystem Interactions with Social Economic Systems, UK Defra Funded Study.
Jiang, Z.J., Huang, X.-P. and Zhang, J.-P. 2010. Effects of CO2 enrichment on photosynthesis, growth, and biochemical composition of seagrass Thalassia hemprichii (Ehrenb.) Aschers. Journal of Integrative Plant Biology 52: 904-913.
Johnson, V.R., Russell, B.D., Fabricius, K.A.E., Brownlee, C. and Hall-Spencer, J.M. 2012. Temperate and tropical brown macroalgae thrive, despite decalcification, along natural CO2 gradients. Global Change Biology 18: 2792-2803.
Keffer, J.E. and Kleinheinz, G.T. 2002. Use of Chlorella vulgaris for CO2 mitigation in a photobioreactor. Journal of Industrial Microbiology and Biotechnology 29: 275-280.
Kleypas, J.A., Anthony, K.R.N. and Gattuso, J.-P. 2011. Coral reefs modify their seawater carbon chemistry -- case study from a barrier reef (Moorea, French Polynesia). Global Change Biology 10.1111/j.1365-2486.2011.02530.x.
Kubler, J.E., Johnston, A.M. and Raven, J.A. 1999. The effects of reduced and elevated CO2 and O2 on the seaweed Lomentaria articulata. Plant, Cell and Environment 22: 1303-1310.
Langdon, C., Broecker, W.S., Hammond, D.E., Glenn, E., Fitzsimmons, K., Nelson, S.G., Peng, T.-S., Hajdas, I. and Bonani, G. 2003. Effect of elevated CO2 on the community metabolism of an experimental coral reef. Global Biogeochemical Cycles 17: 10.1029/2002GB001941.
Leinaas, H.P. and Christie, H. 1996. Effects of removing sea urchins (Strongylocentrotus droebachiensis): stability of the barren state and succession of kelp forest recovery in the east Atlantic. Oecologia 105: 524-536.
Levavasseur, G., Edwards, G.E., Osmond, C.B. and Ramus, J. 1991. Inorganic carbon limitation of photosynthesis in Ulva rotundata (Chlorophyta). Journal of Phycology 27: 667-672.
Ling, S.D., Ibbott, S. and Sanderson, J.C. 2010. Recovery of canopy-forming macroalgae following removal of the enigmatic grazing sea urchin Heliocidaris erythrogramma. Journal of Experimental Marine Biology and Ecology 395: 135-146.
Maberly, S.C. 1990. Exogenous sources of inorganic carbon for photosynthesis by marine macroalgae. Journal of Phycology 26: 439-449.
Manzello, D.P., Enochs, I.C., Melo, N., Gledhill, D.K. and Johns, E.M. 2012. Ocean acidification refugia of the Florida Reef Tract. PLoS ONE 7: e41715.
Martin, G., Torn, K., Blindow, I., Schubert, H., Munsterhjelm, R. and Henricson, C. 2003. Introduction to charophytes. In: Schubert, H. and Blindow, I. (eds.), Charophytes of the Baltic Sea, BMB Publication no. 19, Koeltz Scientific, Königstein, pp. 3-14.
Mercado, J.M., Javier, F., Gordillo, L., Niell, F.X. and Figueroa, F.L. 1999. Effects of different levels of CO2 on photosynthesis and cell components of the red alga Porphyra leucosticia. Journal of Applied Phycology 11: 455-461.
Moulin, L., Catarino, A.I., Claessens, T. and Dubois, P. 2011. Effects of seawater acidification on early development of the intertidal sea urchin Paracentrotus lividus (Lamarck 1816). Marine Pollution Bulletin 62: 48-54.
Muehllehner, N., Glas, M.S. and Lough, J.M. 2011. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nature Climate Change 1: 165-169.
Ow, Y.X., Collier, C.J. and Uthicke, S. 2015. Responses of three tropical seagrass species to CO2 enrichment. Marine Biology 162: 1005-1017.
Pajusalu, L., Martin, G., Pollumae, A., Torn, K. and Paalme, T. 2015. Direct effects of increased CO2 concentrations in seawater on the net primary production of charophytes in a shallow, coastal, brackish-water ecosystem. Boreal Environment Research 20: 413-422.
Palacios, S.L. and Zimmerman, R.C. 2007. Response of eelgrass Zostera marina to CO2 enrichment: possible impacts of climate change and potential for remediation of coastal habitats. Marine Ecology Progress Series 344: 1-13.
Raven, J.A. 2011. Effects on marine algae of changed seawater chemistry with increasing atmospheric CO2. Biology and Environment Proceedings of the Royal Irish Academy 111B: 1-17.
Reiskind, J.B., Seamon, P.T. and Bowes, G. 1988. Alternative methods of photosynthetic carbon assimilation in marine macroalgae. Plant Physiology 87: 686-692.
Ries, J.B., Cohen, A.L. and McCorkle, D.C. 2009. Marine calcifiers exhibit mixed responses to CO2-induced ocean acidification. Geology 37: 1131-1134.
Russell, B.D., Passarelli, C.A. and Connell, S.D. 2011. Forecasted CO2 modifies the influence of light in shaping subtidal habitat. Journal of Phycology 47: 744-752.
Sammarco, P.W., Levington, J.S. and Ogden, J.C. 1974. Grazing and control of coral reef community structure by Diadema antillarum Philippi (Echinodermata: Echinoidea): a preliminary study. Journal of Marine Research 32: 47-53.
Semesi, I.S., Beer, S. and Bjork, M. 2009. Seagrass photosynthesis controls rates of calcification and photosynthesis of calcareous macroalgae in a tropical seagrass meadow. Marine Ecology Progress Series 382: 41-47.
Semesi, I.S., Kangwe, J. and Bjork, M. 2009. Alterations in seawater pH and CO2 affect calcification and photosynthesis in the tropical coralline alga, Hydrolithon sp. (Rhodophyta). Estuarine and Coastal Shelf Science 84: 337-341.
Shan, B.E., Yoshida, Y., Kuroda, E. and Yamashita, U. 1999. Immunomodulating activity of seaweed extract on human lymphocytes in vitro. International Journal of Immunopharmacology 21: 59-70.
Short, F.T., Carruthers, T.J., Dennison, W.C. and Waycott, M. 2007. Global seagrass distribution and diversity: A bioregional model. Journal of Experimental Marine Biology and Ecology 350: 3-20.
Suarez-Alvarez, S., Gomez-Pinchetti, J.L. and Garcia-Reina, G. 2012. Effects of increased CO2 levels on growth, photosynthesis, ammonium uptake and cell composition in the macroalga Hypnea spinella (Gigartinales, Rhodophyta). Journal of Applied Phycology 24: 815-823.
Suetsuna, K. 1998. Separation and identification of angiotensin I-converting enzyme inhibitory peptides from peptic digest of Hizikia fusiformis protein. Nippon Suisan Gakkaishi 64: 862-866.
Surif, M.B. and Raven, J.A. 1989. Exogenous inorganic carbon sources for photosynthesis in seawater by members of the Fucales and the Laminariales (Phaeophyta): ecological and taxonomic implications. Oecologia 78: 97-103.
Torn, K. 2008. Distribution and ecology of charophytes in the Baltic Sea. Ph.D. thesis, University of Tartu.
Troell, M., Halling, C., Neori, A., Chopin, T., Buschmann, A.H., Kautsky, N. and Yarish, C. 2003. Integrated mariculture: asking the right questions. Aquaculture 26: 69-90.
Vogel, N., Fabricius, K.E., Strahl, J., Noonan, S.H.C., Wild, C. and Uthicke, S. 2015. Calcareous green alga Halimeda tolerates ocean acidification conditions at tropical carbon seeps. Limnology and Oceanography 60: 263-275.
Vogel, S. 1996. Life in Moving Fluids: the Physical Biology of Flow. Princeton University Press, Princeton, New Jersey, USA.
Wu, H.-Y., Zou, D.-H. and Gao, K.-S. 2008. Impacts of increased atmospheric CO2 concentration on photosynthesis and growth of micro- and macro-algae. Science in China Series C: Life Sciences 51: 1144-1150.
Xu, Z., Zou, D. and Gao, K. 2010. Effects of elevated CO2 and phosphorus supply on growth, photosynthesis and nutrient uptake in the marine macroalga Gracilaria lemaneiformis (Rhodophyta). Botanica Marina 53: 123-129.
Yang, H., Zhou, Y., Mao, Y., Li, X., Liu, Y. and Zhang, F. 2005. Growth characters and photosynthetic capacity of Gracilaria lemaneiformis as a biofilter in a shellfish farming area in Sanggou Bay, China. Journal of Applied Phycology 17: 199-206.
Yang, Y.F., Fei, X.G., Song, J.M., Hu, H.Y., Wang, G.C. and Chung, I.K. 2006. Growth of Gracilaria lemaneiformis under different cultivation conditions and its effects on nutrient removal in Chinese coastal waters. Aquaculture 254: 248-255.
Zhou, Y., Yang, H., Hu, H., Liu, Y., Mao, Y., Zhou, H., Xu, X. and Zhang, F. 2006. Bioremediation potential of the macroalga Gracilaria lemaneiformis (Rhodophyta) integrated into fed fish culture in coastal waters of north China. Aquaculture 252: 264-276.
Zimmerman, R.C., Kohrs, D.G., Steller, D.L. and Alberte, R.S. 1997. Impacts of CO2-enrichment on productivity and light requirements of eelgrass. Plant Physiology 115: 599-607.
Zou, D. 2005. Effects of elevated atmospheric CO2 on growth, photosynthesis and nitrogen metabolism in the economic brown seaweed, Hizikia fusiforme (Sargassaceae, Phaeophyta). Aquaculture 250: 726-735.
Zou, D., Xia, J. and Yang, Y. 2004. Photosynthetic use of exogenous inorganic carbon in the agarophyte Gracilaria lemaneiformis (Rhodophyta). Aquaculture 237: 421-431.
Last updated 17 November 2015