




Nearly all agricultural species -- including C4 plants -- respond positively to increases in the air's CO2 content by displaying enhanced rates of photosynthesis and biomass production, as well as higher rates of water use efficiency. This summary reviews some of the impacts of these and other related phenomena as they pertain to the C4 crop species of corn (Zea mays L.), or maize as it is often called.
Vanaja et al. (2015) investigated the growth and yield responses of three different maize (Zea mays L.) genotypes -- DHM-117 (a single cross hybrid), Varun (synthetic) and Harsha (composite) -- when exposed to ambient air containing 390 ppm of CO2 or enriched air containing 550 ppm of CO2. As illustrated in the figure below, the nine Indian researchers report that (1) "the improved grain yield due to 550 ppm CO2 was 46% in DHM-117, 61% in Varun and 127% in Harsha as compared with the ambient control," that (2) "the improvement in grain yield was contributed by both increased grain number to the extent of 34%, 25% and 72% as well as enhanced test weight by 8%, 29% and 60% in DHM-117, Varun and Harsha respectively," and that (3) "elevated CO2 also significantly improved the harvest index [Hi] of maize genotypes to the extent of 11% (DHM-117 and Varun) to 68% (Harsha)."
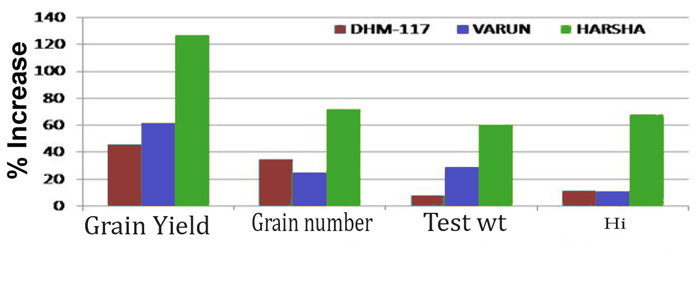
The impact of a 160 ppm increase in CO2 (550 ppm) on various growth and yield parameters of three maize genotypes (DHM-117, Varun and Harsha). Adapted from Vanaja et al. (2015)
As a result of these observations, Vanaja et al. concluded -- in the final sentence of their paper -- that "the positive and significant response of elevated CO2 on maize harvest index was due to higher partitioning of biomass towards reproductive parts rather than vegetative parts," leading them to further conclude that this response "makes this crop more climate resilient."
Maroco et al. (1999) grew maize plants for 30 days in plexiglass chambers maintained at either ambient or triple-ambient concentrations of atmospheric CO2. This exercise revealed that elevated CO2 (1100 ppm) increased maize photosynthetic rates by about 15% relative to those measured in plants grown at 350 ppm CO2, despite of the fact that both rubisco and PEP-carboxylase were down-regulated. This increase in carbon fixation likely contributed to the 20% greater biomass accumulation observed in the CO2-enriched plants. In addition, leaves of CO2-enriched plants contained approximately 10% fewer stomates per unit leaf area than leaves of control plants; and atmospheric CO2 enrichment reduced stomatal conductance by as much as 71% in elevated-CO2-grown plants. As a result of these several different phenomena, the higher atmospheric CO2 concentration greatly increased the intrinsic water-use efficiency of the CO2-enriched plants.
In a study designed to examine the effects of elevated CO2 under real-world field conditions, Leakey et al. (2004) grew maize out-of-doors at the SoyFACE facility in the heart of the United States Corn Belt, while exposing different sections of the field to atmospheric CO2 concentrations of either 354 or 549 ppm. The crop was grown, in the words of the researchers, using cultural practices deemed "typical for this region of Illinois," during a year that turned out to have experienced summer rainfall that was "very close to the 50-year average for this site, indicating that the year was not atypical or a drought year." Then, on five different days during the growing season (11 and 22 July, 9 and 21 August, and 5 September), they measured diurnal patterns of photosynthesis, stomatal conductance and microclimatic conditions.
Contrary to what many people had long assumed would be the case for a C4 crop such as corn growing under even the best of natural conditions, Leakey et al. found that "growth at elevated CO2 significantly increased leaf photosynthetic CO2 uptake rate by up to 41%." The greatest whole-day increase was 21% (11 July) followed by 11% (22 July), during a period of low rainfall. Thereafter, however, during a period of greater rainfall, there were no significant differences between the photosynthetic rates of the plants in the two CO2 treatments, so that over the entire growing season, the CO2-induced increase in leaf photosynthetic rate averaged 10%.
Additionally, on all but the first day of measurements, stomatal conductance was significantly lower (-23% on average) under elevated CO2 compared to ambient CO2, which led to reduced transpiration rates in the CO2-enriched plants on those days as well; and since "low soil water availability and high evaporative demand can both generate water stress and inhibit leaf net CO2 assimilation in C4 plants," they state that the lower stomatal conductance and transpiration rate they observed under elevated CO2 "may have counteracted the development of water stress under elevated CO2 and prevented the inhibition of leaf net CO2 assimilation observed under ambient CO2."
The ultimate implication of their research, in the words of Leakey et al., was that "contrary to expectations, this US Corn Belt summer climate appeared to cause sufficient water stress under ambient CO2 to allow the ameliorating effects of elevated CO2 to significantly enhance leaf net CO2 assimilation." Hence, they concluded that "this response of Z. mays to elevated CO2 indicates the potential for greater future crop biomass and harvestable yield across the US Corn Belt."
In another experiment, Kim et al. (2006) grew well-watered and fertilized maize (Zea mays L. cv. Pioneer 3733) plants from seed to developmental stage R3 (milky ripe stage, 70 days after planting) in sunlit soil-plant-atmosphere research (SPAR) chambers maintained at either 370 (ambient) or 750 (elevated) ppm CO2 concentrations, while periodically measuring a number of plant physiological parameters. They report that at saturating photosynthetically-active radiation (PAR, 2000 µmol m-2 s-1), rates of leaf net photosynthesis in the elevated CO2 chambers were only 4% greater than those in the ambient CO2 chambers. However, upper-canopy leaves grown at elevated CO2 exhibited reductions in excess of 50% in both leaf stomatal conductance and transpiration; and as a result of these changes, instantaneous leaf water use efficiency was more than doubled in the high-CO2 treatment. Because of shading within the canopy, however, and because the difference in stomatal conductance between ambient and elevated CO2 declines as PAR drops, the CO2-induced reduction in canopy evapotranspiration was only a little over 20%. Consequently, although the maize plants of this study were somewhat anomalous in their failure to significantly boost their growth in response to atmospheric CO2 enrichment, their more characteristic transpiration response enabled them to produce their slightly enhanced biomass with a considerably smaller consumptive use of water than that of the plants growing in ambient air.
Chun et al. (2011) grew corn plants from seed in naturally-sunlit soil-plant-atmosphere-research (SPAR) units in which temperature, humidity and CO2 concentration were precisely controlled, the latter at either 400 ppm (ambient) or 800 ppm (elevated), beginning 21 days after emergence (DAE). These units were placed atop soil bins (2.0 m long by 0.5 m wide by 1.0 m deep) that were filled with a mixture of 75% coarse sand and 25% vermiculate, where soil water contents were monitored hourly by a time domain reflectometry (TDR) system that consisted of 15 TDR probes per chamber placed within three rows at depths of 0, 15, 30, 50 and 75 cm from the soil surface. By means of this system of soil water content assessment, combined with nightly "fertigation," Chun et al. were able to provide the plants with the nitrogen they needed while maintaining four different soil water stress levels -- control, mild, moderate and severe -- which were also initiated 21 DAE. Thereafter, the height, number of leaves, leaf lengths and growth states of the corn plants were determined twice weekly, while samples of the plants were collected, dried and analyzed for biomass accumulation at 21 and 60 DAE (the beginning and the end of the different CO2 and soil water content treatments).
In discussing their findings, the five researchers report that under both well-watered and water-stressed conditions, higher soil water contents were maintained in the elevated CO2 treatment, even though 20-49% less water was applied to the soil of the elevated CO2 treatment. Their study did not, however, provide any evidence that the elevated CO2 treatment had a strong effect on plant height, leaf area or above-ground biomass. But the water saving was amazing; and as a result they concluded, that "under increased CO2 concentrations as generally predicted in the future, less water will be required for corn plants than at present." And since water is already a scarce commodity in many parts of the world -- and will only become scarcer, more expensive and more difficult to obtain in the days and years ahead -- this finding is extremely welcome news.
Writing as background for their work, Allen et al. (2011) state that "plants of the C4 photosynthetic pathway have a CO2-concentrating mechanism that overcomes limitations of low atmospheric CO2" and which thereby provides them with "a near-saturating photosynthetic capability at current atmospheric CO2." In this circumstance, as they continue, "a rise in atmospheric CO2 will theoretically have a limited direct impact on C4 photosynthesis." Nevertheless, they note that "a number of C4 crop plants express a positive response to elevated growth CO2, although to a smaller extent compared to C3 plants," citing the analyses of Kimball (1993) and Poorter et al. (1996).
As their contribution to the subject, Allen et al. grew maize (Zea mays L. cv. Saturn Yellow) and grain sorghum (Sorghum bicolor L. cv. DeKalb 28E) seeds for 39 days in sunlit controlled-environment chambers at 360 and 720 ppm CO2 concentrations, while throughout this period canopy net photosynthesis and evapotranspiration were measured and summarized daily from 08:00 to 17:00 hours, with irrigation being withheld from matched pairs of treatments starting 26 days after sowing, and with biomass determinations being made at 34 and 39 days after sowing for maize and grain sorghum, respectively.
The four researchers report that their data indicated that for both maize and grain sorghum, there was a "maintenance of relatively high canopy photosynthetic rates in the face of decreased transpiration rates [that] resulted in enhanced water use efficiency when these plants were grown at elevated CO2 of 720 ppm, but not at 360 ppm." And as a result, they demonstrated that "both plants maintained growth better at double-ambient CO2 than their counterparts at ambient CO2 in the presence of drought," such that reductions in total above-ground biomass due to drought were 42% for maize and 36% for sorghum at ambient CO2, but only 18% for maize and 14% for sorghum at double-ambient CO2.
In discussing their several findings, Allen et al. write in their paper's last paragraph that they "agree with Leakey (2009) that drought stress in C4 crop plants can be ameliorated at elevated CO2 as a result of lower stomatal conductance and sustained intercellular CO2." And as a result of this experimentally-observed fact, they conclude that "management of irrigation water in a future high CO2 world could potentially increase overall C4 crop yields (in water-limited areas) by allocating limited water supplies [so as to be able] to irrigate larger crop areas at sub-optimal [as opposed to conventional optimal] plant-available soil water levels, at least during vegetative stages of growth," thereby obtaining greater total yields with the same allocation of available water.
Also focusing on drought, Meibaum et al. (2012) write that maize (a C4 crop) "is one of the most important feedstuffs for ruminants but studies investigating the combined effects of drought and CO2 elevation on the plant and its fermentation in the rumen are scarce," citing Loholter et al. (2012); and they thus state that "there is an urgent need to investigate the effect of climate change, i.e., increase of atmospheric CO2 and of summer drought on rumen fermentation in a more detailed way."
Against this backdrop, in a FACE study conducted at an experimental field of the Federal Research Institute for Rural Areas, Forestry and Fisheries, which is located in Braunschweig (Germany), Meibaum et al. grew maize (Zea mays L., cv. Romario) at atmospheric CO2 concentrations of either 380 or 550 ppm, and at either 170 or 330 mm of total growing-season water supply. Then, at the end of the growing season, whole maize plants were harvested and representative samples of each of them were analyzed for crude nutrients, neutral detergent fiber, acid detergent fiber and acid detergent lignin (ADL). In addition, in an in vitro experiment using the semi-continuous rumen-simulation technique (RUSITEC), as originally described by Czerkawski and Breckenridge (1977), they studied the effects of atmospheric CO2 enrichment on rumen fermentation patterns.
The six German researchers report that "the Bacteria- and Archaea-specific SSCP [single strand conformation polymorphism] profiles as well as the production rates of short-chain fatty acids and their molar percentages were not affected by [the various] treatments." They also state that "the digestibilities of organic matter obtained from the present experiment (45%-48%) are consistent with the results from RUSITEC experiments carried out by other authors (Czerkawski, 1986; Klevenhusen et al., 2009; Hildebrand et al., 2010)." And they additionally indicate that "the CO2 treatment had no effect on digestibility of organic matter, which is in agreement with the findings of Akin et al. (1994), who studied the effect of elevated CO2 on sudan grass," which is another C4 plant. Meibaum et al. thus conclude that "changes in forage due to climate change will not result in significant alterations of rumen microbial community because of its pronounced ability of dynamic adaptation," and, therefore, they suggest that "most parameters of rumen digestion will only be slightly influenced by plant material altered as a result of climatic changes."
Studying the combined effects of nitrogen stress and drought, Zong and Shangguan (2014) grew maize (Zea mays L. cv. Zhengdan 958) seedlings hydroponically cultured in sand within two climate-controlled chambers with CO2 concentrations of either 380 or 750 ppm and N solutions of either 5 mM N (N deficient treatment) and 15 mM N (control N treatment. The two Chinese scientists report that "maize seedlings suffering combined N limitation and drought had a better recovery of new leaf photosynthetic potential than those suffering only drought with ambient CO2." But they say that with elevated CO2, "the plants were able to maintain favorable water content as well as enhance their biomass accumulation, photochemistry activity, leaf water use efficiency and new leaf growth recoveries." As a result, Zong and Shangguan conclude that "elevated CO2 could help drought-stressed seedlings to maintain higher carbon assimilation rates under low water content," noting that such was the case "even under N-limited conditions, which allow the plants to have a better performance under drought following re-watering."
Also germane to this subject and supportive of the above conclusion are the effects of elevated CO2 on weeds associated with corn. Conway and Toenniessen (2003), for example, report on maize in Africa being attacked by the parasitic weed Striga hermonthica, which sucks vital nutrients from its roots, as well as from the roots of many other C4 crops of the semi-arid tropics, including sorghum, sugar cane and millet, plus the C3 crop rice, particularly throughout much of Africa, where Striga is one of the region's most economically important parasitic weeds. However, other researchers have shown that atmospheric CO2 enrichment greatly reduces the damage done by this devastating weed [see, for example, Watling and Press (1997) and Watling and Press (2000)].
Baczek-Kwinta and Koscielniak (2003) studied yet another phenomenon that is impacted by atmospheric CO2 enrichment and that can affect the productivity of maize. Noting the tropical origin of maize and that the crop "is extremely sensitive to chill (temperatures 0-15°C)," they report that it is nevertheless often grown in cooler temperate zones because of its high yield potential. In such circumstances, however, maize can experience a variety of maladies associated with exposure to periods of low air temperature. Hence, to see if elevated CO2 either exacerbates or ameliorates this problem, they grew two hybrid genotypes -- KOC 9431 (chill-resistant) and K103xK85 (chill-sensitive) -- from seed in air of either ambient (350 ppm) or elevated (700 ppm) CO2 concentration (AC or EC, respectively), after which they exposed the plants to air of 7°C for eleven days, whereupon they let them recover for one day in ambient air of 20°C, all the while measuring several physiological and biochemical parameters pertaining to the plants' third fully-expanded leaves.
The two researchers' protocol revealed that "EC inhibited chill-induced depression of net photosynthetic rate (PN), especially in leaves of chill-resistant genotype KOC 9431," which phenomenon "was distinct not only during chilling, but also during the recovery of plants at 20°C." In fact, they found that "seedlings subjected to EC showed 4-fold higher PN when compared to AC plants." They also determined that "EC diminished the rate of superoxide radical formation in leaves in comparison to the AC control." In addition, they found that leaf membrane injury "was significantly lower in samples of plants subjected to EC than AC." Last of all, they report that enrichment of the air with CO2 successfully inhibited the decrease in the maximal quantum efficiency of photosystem 2, both after chilling and during the one-day recovery period. And in light of all of these positive effects of elevated CO2, they concluded that "the increase in atmospheric CO2 concentration seems to be one of the protective factors for maize grown in cold temperate regions."
But what about the effects of climate change, both past and possibly future, on corn production? For nine areas of contrasting environment within the Pampas region of Argentina, Magrin et al. (2005) evaluated changes in climate over the 20th century along with changes in the yields of the region's chief crops. Then, after determining upward low-frequency trends in yield due to technological improvements in crop genetics and management techniques, plus the aerial fertilization effect of the historical increase in the air's CO2 concentration, annual yield anomalies and concomitant climatic anomalies were calculated and used to develop relations describing the effects of changes in precipitation, temperature and solar radiation on crop yields, so that the effects of long-term changes in these climatic parameters on Argentina agriculture could be determined.
Noting that "technological improvements account for most of the observed changes in crop yields during the second part of the 20th century," which totaled 110% for maize, Magrin et al. report that due to changes in climate between the periods 1950-70 and 1970-99, maize yields increased by 18%. As a result, it can be appreciated that late 20th-century climate change, which is claimed by climate alarmists to have been unprecedented over the past two millennia and is often described by them as one of the greatest threats ever to be faced by humanity, has definitely not been a problem for corn cultivation in Argentina. In fact, it has actually helped it.
Much the same has been found to be true in Alberta, Canada, where Shen et al. (2005) derived and analyzed long-term (1901-2002) temporal trends in the agroclimate of the region. They report, for example, that "an earlier last spring frost, a later first fall frost, and a longer frost-free period are obvious all over the province." They also found that May-August precipitation in Alberta increased 14% from 1901 to 2002, and that annual precipitation exhibited a similar increasing trend, with most of the increase coming in the form of low-intensity events. In addition, the researchers note that "the area with sufficient corn heat units for corn production, calculated according to the 1973-2002 normal, has extended to the north by about 200-300 km, when compared with the 1913-32 normal, and by about 50-100 km, when compared with the 1943-72 normal."
In light of these findings, Shen et al. conclude that "the changes of the agroclimatic parameters imply that Alberta agriculture has benefited from the last century's climate change," emphasizing that "the potential exists to grow crops and raise livestock in more regions of Alberta than was possible in the past." They also note that the increase in the length of the frost-free period "can greatly reduce the frost risks to crops and bring economic benefits to Alberta agricultural producers," and that the northward extension of the corn heat unit boundary that is sufficient for corn production "implies that Alberta farmers now have a larger variety of crops to choose from than were available previously." Hence, they say "there is no hesitation for us to conclude that the warming climate and increased precipitation benefit agriculture in Alberta."
Writing as background for their work, Chen et al. (2011) state that corn acreage in Northeast China -- comprised of Heilongjiang, Jilin and Liaoning provinces -- "accounts for 26.3% of the corn area in the country and accounts for about 29.4% of Chinese total corn grain production." And in light of these facts, they say that the corn production of this region "plays a significant role in ensuring Chinese food security," noting that "knowledge of the potential effects of climate change on corn production in Northeast China will be highly valuable, not only for China but also for the world."
To obtain this important knowledge, Chen et al. "performed a multiple regression analysis to study the relationship between corn growth and the key climatic factors of temperature and precipitation during the crop growing season and during specific months in Northeast China from 1965 to 2008," in order to learn "which variables of climate change and which month in the crop growing season were associated with yield variability over this 44-year period." In so doing, the six scientists determined that the major climate factor affecting corn yield in Northeast China is daily minimum temperature, particularly in the months of May and September; and they found that a warming of 1.0°C in the mean daily minimum temperature of either of these months may enhance corn yield by either 303 kg/ha or 284 kg/ha, respectively. In addition, they found that growth duration -- defined as the period from sowing to harvest (days to maturity) -- rose by six days in Liaoning province and by seven days in Jilin and Heilongjiang provinces over the period 1950-2008, which encompassed the period of 1965-2008, when daily minimum temperature rose at a rate of 0.44°C per decade. And in light of such findings, Chen et al. conclude that "in order to fully exploit the positive effects of global warming on corn production, new varieties should be adapted to the longer growing season," which adjustment could be described as a good example of man and nature working together to insure maximum food security for a large portion of mankind.
With respect to the future, Bootsma et al. (2005) derived relationships between agroclimatic indices and average yields of major grain crops, including corn, from field trials conducted in eastern Canada, after which they used them to estimate potential impacts of projected climate change scenarios on anticipated average yields for the period 2040 to 2069. Based on a range of available heat units projected by multiple General Circulation Model (GCM) experiments, they determined that average yields achievable in field trials could increase by 40 to 115% for corn, "not including the direct effect of increased atmospheric CO2 concentrations." Adding expected CO2 increases to the mix, along with gains in yield anticipated to be achieved through breeding and improved technology, these numbers rose to 114 to 186%.
In light of their fantastic findings, Bootsma et al. predict there will be a "switch to high-energy and high-protein-content crops (corn and soybeans) that are better adapted to the warmer climate." Consequently, if the GCM-based climate-change predictions prove correct, Canada will be immensely blessed by the incredible boost the changed conditions will bring to the country's agricultural productivity, and especially that of corn.
In yet another study, based on what is known about the effects of rising temperatures and atmospheric CO2 concentrations on plant growth and development, as well as what might be expected to occur in terms of yet to come technological innovations, Costa et al. (2009) estimated the impacts of these three phenomena on the yields of maize (Zea mays) and common beans (Phaseolus vulgaris) in Brazil, where the degree of global warming they investigated is that which is projected for the years 2050 and 2080 by the HadCM3 climate model when it is driven by the greenhouse gas concentration increases that are specified in the A2 scenario of the IPCC's Special Report on Emissions Scenarios.
Under this scenario, Costa et al. calculate that the warming conditions associated with increased greenhouse gases "lead to reductions in the potential productivity of maize and beans for the years 2050 and 2080 by up to 30%." However, they say that the CO2 fertilization effect is expected to overcome the negative response to warming and lead to a net increase in the productivity of common beans. In the case of maize, on the other hand, they find that "the CO2 fertilization feedback is much weaker and cannot cancel out the thermal effect." But when they include ever-evolving technology in the mix, they find that "appropriate soil and technological management as well as genetic improvements may very likely induce an increase in bean and maize yield despite the unfavorable future climate conditions."
Once again, human ingenuity -- aided by just one of the many positive biological effects of the ongoing rise in the air's CO2 content -- would appear to be capable of enabling the agricultural enterprise to continue to meet the world's food needs, even in the face of the unrealistically high temperature increases predicted by the IPCC to occur over the next several decades.
An optimistic future for maize yields is also gleaned from the study of Butler and Huybers (2013), who write as background for their work that "high temperatures are associated with reduced crop yields, and predictions for future warming have raised concerns regarding future productivity and food security." More specifically, they note that "global maize yields are forecast to decline in response to increasing temperature, particularly as the upper range of growing season temperatures becomes hotter," citing Schlenker and Roberts (2006), Easterling et al. (2007), Lobell and Field (2007), Battisti (2009), Lobell et al. (2011) and Roberts and Schlenker (2011). However, they caution that "the extent to which adaptation can mitigate such heat-related losses remains unclear," and they thus proceed to present some much-needed clarity (and optimism) on this subject.
In their study, Butler and Huybers first proceed to empirically demonstrate how maize is locally adapted to hot temperatures across a subset of 1,013 US counties, after which they used this spatial adaptation "as a surrogate for future adaptation," noting that "US corn hybrids have a product half-life of about 4 years, suggesting sufficiently rapid turnover to adapt to decadal changes in climate." And with the help of this adaptation phenomenon, the two researchers were able to determine that (1) "losses to average US maize yields from a 2°C warming would be reduced from 14% to only 6%," and that (2) "loss in net production is wholly averted."
As for a few specifics on the matter, they indicate that under the 2°C warming scenario, "Minnesota stands to increase yields by 11%; the yield losses from northern Ohio west to northern Missouri are nearly eliminated; and North Carolina, Georgia and east Texas reduce losses from 49% without adaptation to 39% with it." Given as much, in the concluding paragraph of their report, Butler and Huybers write that "losses to US maize yield from increased temperature," such as those suggested by Schlenker and Roberts (2006, 2009), "are almost certainly overestimated if adaptation is not accounted for." But if it is a part of the analysis, their work suggests that there could well be no net loss in productivity across the entire corn-production region.
In summary, it seems fairly clear that as the air's CO2 content continues to rise, and even if the climate of the world changes in the ways suggested by GCM calculations, maize plants will likely display greater rates of photosynthesis and biomass production, as well as reduced transpirational water losses and increased water-use efficiencies; and to top it all off, more areas of the world will likely become suitable for growing this important crop.
For an even larger listing of such responses, see the Plant Growth Data section of the CO2 Science website and view the Dry Weight and Photosynthesis responses for maize under atmospheric CO2 enrichment listed there.
References
Akin, D.E., Kimball, B.A., Mauney, J.R., LaMorte, R.L., Hendrey, G.R., Lewin, K., Nagy, J. and Gates, R.N. 1994. Influence of enhanced CO2 concentration and irrigation on sudan grass digestibility. Agricultural and Forest Meteorology 70: 279-287.
Allen Jr., L.H., Kakani, V.G., Vu, J.C.V. and Boote, K.J. 2011. Elevated CO2 increases water use efficiency by sustaining photosynthesis of water-limited maize and sorghum. Journal of Plant Physiology 168: 1909-1918.
Baczek-Kwinta, R. and Koscielniak, J. 2003. Anti-oxidative effect of elevated CO2 concentration in the air on maize hybrids subjected to severe chill. Photosynthetica 41: 161-165.
Battisti, D.S. and Naylor, R.L. 2009. Historical warnings of future food insecurity with unprecedented seasonal heat. Science 323: 240-244.
Bootsma, A., Gameda, S. and McKenney, D.W. 2005. Potential impacts of climate change on corn, soybeans and barley yields in Atlantic Canada. Canadian Journal of Plant Science 85: 345-357.
Butler, E.E. and Huybers, P. 2013. Adaptation of US maize to temperature variations. Nature Climate Change 3: 68-72.
Chen, C., Lei, C., Deng, A., Qian, C., Hoogmoed, W. and Zhang, W. 2011. Will higher minimum temperatures increase corn production in northeast China? An analysis of historical data over 1965-2008. Agricultural and Forest Meteorology 151: 1580-1588.
Chun, J.A., Wang, Q., Timlin, D., Fleisher, D. and Reddy, V.R. 2011. Effect of elevated carbon dioxide and water stress on gas exchange and water use efficiency in corn. Agricultural and Forest Meteorology 151: 378-384.
Conway, G. and Toenniessen, G. 2003. Science for African food security. Science 299: 1187-1188.
Costa, L.C., Justino, F., Oliveira, L.J.C., Sediyama, G.C., Ferreira, W.P.M . and Lemos, C.F. 2009. Potential forcing of CO2, technology and climate changes in maize (Zea mays) and bean (Phaseolus vulgaris) yield in southeast Brazil. Environmental Research Letters 4: 10.1088/1748-9326/4/1/014013.
Czerkawski, J.W. 1986. An Introduction to Rumen Studies. Pergamon Press Ltd., Oxford, United Kingdom.
Czerkawski, J.W. and Breckenridge, G. 1977. Design and development of a long-term rumen simulation technique (Rusitec). British Journal of Nutrition 38: 371-384.
Easterling, W., Aggarwal, P., Batima, P., Brander, K., Erda, L., Howden, M., Kirilenko, A., Morton, J., Soussana, J.-F., Schmidhuber, S. and Tubiello, F. 2007. Food, fibre and forest products. In: Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J. and Hanson, C.E. (Eds.). Cambridge University Press, Cambridge, United Kingdom, pp. 273-313.
Hildebrand, B., Boguhn, J. and Rodehutscord, M. 2010. Effect of maize silage to grass silage ratio and feed particle size on ruminal fermentation in vitro. Animal Feed Science and Technology 5: 528-536.
Kim, S.-H., Sicher, R.C., Bae, H., Gitz, D.C., Baker, J.T., Timlin, D.J. and Reddy, V.R. 2006. Canopy photosynthesis, evapotranspiration, leaf nitrogen, and transcription profiles of maize in response to CO2 enrichment. Global Change Biology 12: 588-600.
Kimball, B.A. 1993. Effects of elevated CO2 and climate variables on plants. Journal of Soil and Water Conservation 48: 9-14.
Klevenhusen, F., Bernasconi, S.M., Hofstetter, T.B., Bolotin, J., Kunz, C. and Soliva, C.R. 2009. Efficiency of monolaurin in mitigating ruminal methanogenesis and modifying C-isotope fraction when incubating diets composed of either C3 or C4 plants in a rumen simulation technique (Rusitec) system. British Journal of Nutrition 102: 1308-1317.
Leakey, A.D.B. 2009. Rising atmospheric carbon dioxide concentration and the future of C4 crops for food and fuel. Proceedings of the Royal Society B 276: 2333-2343.
Leakey, A.D.B., Bernacchi, C.J., Dohleman, F.G., Ort, D.R. and Long, S.P. 2004. Will photosynthesis of maize (Zea mays) in the US Corn Belt increase in future [CO2] rich atmospheres? An analysis of diurnal courses of CO2 uptake under free-air concentration enrichment (FACE). Global Change Biology 10: 951-962.
Lobell, D.B., Banziger, M., Magorokosho, C. and Vivek, B. 2011. Nonlinear heat effects on African maize as evidenced by historical yield trials. Nature Climate Change 1: 42-45.
Lobell, D.B. and Field, C.B. 2007. Global scale climate-crop yield relationships and the impacts of recent warming. Environmental Research Letters 2: 10.1088/1748-9326/2/1/014002.
Loholter, M., Meyer, U., Manderscheid, R., Weigel, H.J., Erbs, M., Flachowsky, G. and Danicke, S. 2012. Effects of free air carbon dioxide enrichment and drought stress on the feed value of maize silage fed to sheep at different thermal regimes. Archives of Animal Nutrition 66: 335-346.
Magrin, G.O., Travasso, M.I. and Rodriguez, G.R. 2005. Changes in climate and crop production during the 20th century in Argentina. Climatic Change 72: 229-249.
Maroco, J.P., Edwards, G.E. and Ku, M.S.B. 1999. Photosynthetic acclimation of maize to growth under elevated levels of carbon dioxide. Planta 210: 115-125.
Meibaum, B., Riede, S., Schroder, B., Manderscheid, R., Weigel, H.-J. and Breves, G. 2012. Elevated CO2 and drought stress effects on the chemical composition of maize plants, their ruminal fermentation and microbial diversity in vitro. Archives of Animal Nutrition 66: 473-489.
Poorter, H., Roumet, C. and Campbell, B.D. 1996. Interspecific variation in the growth response of plants to elevated CO2: a search for functional types. In: Korner, C. and Bazzaz, F.A. (Eds.). Carbon Dioxide, Populations, and Communities. Academic Press, New York, New York, USA, pl 375-412.
Roberts, M. and Schlenker, W. 2011. In: Libecap, G. and Steckel, R. (Eds.) The Economics of Climate Change, Adaptations Past and Present. University of Chicago Press, Chicago, Illinois, USA, pp. 225-252.
Schlenker, W. and Roberts, M.J. 2006. Nonlinear effects of weather on corn yields. Review of Agricultural Economics 28: 391-398.
Schlenker, W. and Roberts, M.J. 2009. Nonlinear temperature effects indicate severe damage to U.S. crop yields under climate change. Proceedings of the National Academies of Science USA 106: 15,594-15,598.
Shen, S.S.P., Yin, H., Cannon, K., Howard, A., Chetner, S. and Karl, T.R. 2005. Temporal and spatial changes of the agroclimate in Alberta, Canada, from 1901 to 2002. Journal of Applied Meteorology 44: 1090-1105.
Watling, J.R. and Press, M.C. 1997. How is the relationship between the C4 cereal Sorghum bicolor and the C3 root hemi-parasites Striga hermonthica and Striga asiatica affected by elevated CO2? Plant, Cell and Environment 20: 1292-1300.
Watling, J.R. and Press, M.C. 2000. Infection with the parasitic angiosperm Striga hermonthica influences the response of the C3 cereal Oryza sativa to elevated CO2. Global Change Biology 6: 919-930.
Zong, Y. and Shangguan, Z. 2014. CO2 enrichment improves recovery of growth and photosynthesis from drought and nitrogen stress in maize. Pakistan Journal of Botany 46: 407-415.
Last updated 29 December 2015