




Most of the ocean acidification research conducted to date has focused solely on the biological impacts of declining seawater pH. Fewer studies have investigated the interactive effects of ocean acidification and temperature. This summary examines what has been learned in several of such studies for coral reefs, as reported in various field-based studies on the topic. Contrary to what is widely assumed and reported, the studies reviewed here collectively reveal that many corals will remain unaffected by rising temperatures and atmospheric CO2 concentrations. Furthermore, in contrast to projections, some will likely experience growth and performance benefits.
According to Cantin et al. (2009), "zooxanthellae (symbiotic dinoflagellates of the genus Symbiodinium) are critical to the survival of reef-building corals, providing a major source of energy from photosynthesis for cell maintenance, growth and reproduction of their coral hosts," which services, include the energetically-expensive process of calcification (Idso et al., 2000). Against this backdrop, Cantin et al. studied the amount of photosynthetic "rent" paid by two different clades of Symbiodinium (C1 and D) to their coral hosts (juvenile Acropora millepora) for the privilege of living within the latter's calcareous "houses." This was done by measuring the "financial transfer" to nine-month-old corals that had been developed "from crosses involving the same parent corals," which "planned parenthood" minimized any host genetic differences that might otherwise have influenced the physiology of the host-symbiont "lease agreement."
Results indicated that "Symbiodinium C1 exhibited a 121% greater capacity for translocation of photosynthate to A. millepora juveniles along with 87% greater relative electron transport through photosystem II under identical environmental conditions." In addition, the five researchers note that "A. tenuis and A. millepora juveniles in a previous study exhibited 2 to 3 times faster growth rates when associated with Symbiodinium C1 compared to those associated with Symbiodinium D (Little et al., 2004) at the same field site where juveniles were reared in the present study."
As a result of their analysis, Cantin et al. conclude that "the differences in carbon-based energy transfer between symbiont types may provide a competitive advantage to corals associating with Symbiodinium C1, particularly during their early life histories, when greater energy investment into rapid tissue and skeletal growth can prevent overgrowth of juveniles by competitors and mortality from grazers." Likewise, they opine that "as the community structure of coral reefs shift in response to global climate change [global warming?] and water quality impacts [ocean acidification?], opportunistic corals harboring symbionts that enable maximum rates of growth may similarly gain a competitive advantage." Consequently, it would appear that in the economy of nature ample provision has been made for Earth's corals to weather all sorts of environmental challenges that may come their way, including those that the IPCC contends will be driven by rising atmospheric CO2 concentrations.
In studying coral calcification rates on Australia's Great Barrier Reef, De'ath et al. (2009) report there has been a 14% drop in Porites calcification rate from 1990 to 2005 (although a graphical view of their data indicate the decline began around 1970) and that this decline "is unprecedented in at least the past 400 years," which is indeed what their data show. But, such statements do not reveal the full story. If, for example, their calcification history is followed back in time a mere 33 more years, from 1605 to 1572 -- when the air's CO2 concentration was more than 100 ppm less than what it is today and, therefore, was supposedly so much more healthier for corals (according to the ocean acidification hypothesis) -- the coral calcification rate at that earlier time is seen to have been approximately 21% lower than what it was at its 20th-century peak.
Another way of looking at De'ath et al.'s data is to realize that from 1572 to 1970, Porites calcification rates on the Great Barrier Reef rose, by about 27%, as the atmospheric CO2 concentration and air temperature rose concurrently, after which calcification rates declined, but by a much smaller 14%, as these same air temperature and CO2 trends continued, further obfuscating the issue. Perhaps recognizing this dichotomy, De'ath et al. add that "the causes for the [1990 to 2005] Great Barrier Reef-wide decline in coral calcification of massive Porites remain unknown." Three years later, however, the work of another research team provided a possible answer.
Introducing their work, Uthicke et al. (2012) state that "tropical coral reefs are currently under threat by a variety of regional and global stressors," with examples of the former being "land runoff and overfishing (e.g., Pandolfi et al., 2003; Fabricius, 2005)," while examples of the latter are "sea temperature increase and ocean acidification (Hoegh-Guldberg et al., 2007; Fabricius et al., 2011)." In an effort designed to determine which set of stressors is the most significant or bigger threat, Uthicke et al. analyzed sediment cores collected from inshore fringing coral reefs in the Whitsunday area of the Great Barrier Reef (GBR) of Australia. These cores were collected from three different locations: (1) inner near-shore reefs with low coral cover and high macroalgal abundance, (2) intermediate reefs, and (3) reefs at outer islands with low algal and high coral cover. These three locations were chosen based on the facts that (a) inner near-shore reefs are typically the first to produce evidence of regional human impacts, that (b) reefs at outer islands are the last to experience the negative effects of human influence, as well as the most likely to exhibit evidence of global stressors, while (c) intermediate reefs often show evidence of both.
The three researchers report their results indicated that benthic foraminiferal assemblages found in the cores of outer-island reefs that are unaffected by increased land runoff have been "naturally highly persistent over long (>2000 years) timescales." In both of the other zones, assemblages were also rather persistent, but only until 150 years ago; and they say that assemblages <55 years old from inner near-shore and intermediate reefs were significantly different from older assemblages.
In concluding their report, Uthicke et al. write that they found support for the likelihood that "increased land runoff since the start of land clearing and agriculture in the catchment of the Whitsunday Region of the GBR has left a signature in the foraminiferal assemblages of inner and intermediate areas of the study area," when previously the assemblages of these areas had been "persistent for at least several thousand years." In addition, and based on the fact that "no changes were observed on outer reefs located away from land runoff," they propose that "changes observed on inner and intermediate reefs were mainly driven by enhanced agricultural runoff after European settlement." And topping off everything else, they affirm that "the hypothesis that global forcing, such as sea temperature increase or ocean acidification, altered the foraminiferal community found little support." In fact, it found none.
Also working on the Great Barrier Reef was Browne (2012), who writes that "local stressors erode reef resilience, and therefore increase their vulnerability to global stressors that include ocean warming, [which is] predicted to increase the severity and intensity of coral bleaching events (Hoegh-Guldberg, 1999), ocean acidification, [which is] predicted to reduce calcification rates and reef growth (Kleypas et al., 1999), and increased storm and cyclone activity, [which is] predicted to reduce coral framework complexity and stability (Puotinen, 2004)." In a study designed to assess the overall impact of these several reef stressors, Browne describes how "coral growth rates (linear extension, density, calcification rates) of three fast-growing corals (Acropora, Montipora, Turbinaria) were studied in situ on Middle Reef, an inshore reef located on the central Great Barrier Reef (GBR)," in order to "assess the influence of changing environmental conditions on coral condition and reef growth."
Results indicate that "despite local anthropogenic pressures and global climate change, Middle Reef has a robust and resilient coral community," noting that "Acropora linear extension rates were comparable with rates observed at similar depths and sea surface temperatures on mid to offshore reefs on the GBR, and in the Caribbean," while additionally indicating that "Montipora and Turbinaria are abundant on inshore turbid reefs due to their adaptive capacities and are therefore an important source of carbonate for reef growth and development." In fact, Browne writes that "Montipora linear extension was greater than current estimates available, and Turbinaria, although characterized by slow linear extension, had a dense skeleton and hence may be more resilient to physical damage as ocean pH falls." And of both of them, Browne states that although they "may be more susceptible during the warmer months due to multiple stressors, they were able to rapidly recover during the cooler months." "In summary," as Browne concludes, "corals on Middle Reef are robust and resilient to their marginal environmental conditions."
Helmle et al. (2011) collected coral cores in May of 1997 and June of 1998 from seven Montastraea faveolata colonies located in the upper Florida Keys (USA), where they "were drilled at the location of maximum vertical growth of the colony." Chronologies were constructed from the annual density bands found in the cores, and it was determined that all of them had a 60-year common period from 1937 to 1996. And for these cores, annual extension, density and calcification rates were obtained and analyzed, to see how they varied over this period of intensifying warming and acidification of the global ocean.
The five U.S. scientists report that their data show "no evidence of significant age effects" over the 1937-1996 period for either extension, density or calcification. In discussing their findings, Helmle et al. say they demonstrate that "the measured corals have historically been able to maintain rates of extension and calcification over the 60-year period from 1937 to 1996 under the combination of local environmental and climatic changes." They also note that calcification rates were positively related to sea surface temperature, "similar to results for Porites corals from Tahiti (Bessat and Buigues, 2001) and the Great Barrier Reef (Lough and Barnes, 1997)," but they say that the Florida results only explained about 7% of the annual calcification variability as opposed to ~30% at the Pacific locations.
As for why the Florida Keys corals have fared so well, Helmle et al. write that the answer could be that "massive reef-building corals are not as susceptible to declines in Ωarag [aragonite saturation state] as demonstrated by laboratory experiments; local processes, such as high seasonal variation in Ωarag in the Florida Keys, may be temporarily enabling these corals to maintain their historical rates of calcification; the role of Ωarag in controlling calcification is masked amidst considerable natural inter-annual variability; or the actual in situ reef-site carbonate chemistry is decoupled from the oceanic values, which could occur as a result of shifts in benthic community metabolism, mineral buffering and/or coastal biogeochemical processes."
In another analysis, Shi et al. (2012) state that "rising atmospheric CO2 and global warming are regarded as fatal threats to coral reefs," noting that "the IPCC has reported that by the end of this century, coral reefs will be the first ecological system that will become extinct," citing Wilkinson (2004). However, they say "others contend that rising seawater temperature is conducive to enhanced coral calcification, and increased calcification will be higher than the decline caused by rising CO2," so that "coral calcification will increase by about 35% beyond pre-industrial levels by 2100, and no extinction of coral reefs will occur in the future," citing McNeil et al. (2004).
In an attempt to shed some light on these contrasting viewpoints, in late May of 2004 and 2007 Shi et al. extracted core samples of coral skeletons from several massive live and dead Porites lutea colonies comprising part of the Meiji Reef in the southern South China Sea, after which they analyzed their skeletal calcification rates by means of X-ray photography, which enabled them to construct a nearly three-century-long history of coral calcification rate for the period 1716-2005. The results of the six scientists' efforts are depicted in Figure 1.
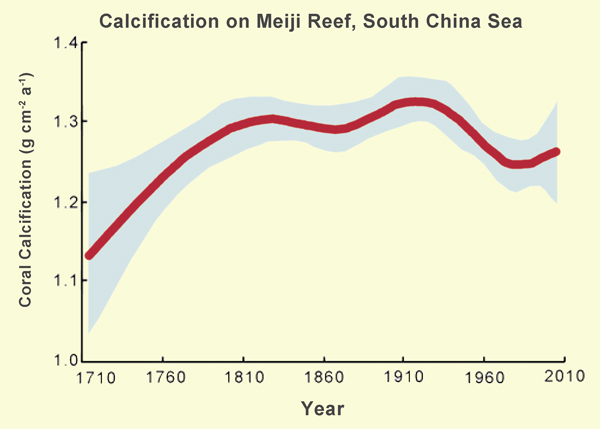
Figure 1. The long-term history of coral calcification rate on Meiji Reef. Adapted from Shi et al. (2012).
As best as can be determined from the Chinese scientists' graph, over the period of time depicted -- when the IPCC claims the world warmed at a rate that was unprecedented over the past millennium or two, and when the atmosphere's CO2 concentration rose to values not seen for millions of years -- the two "fatal threats to coral reefs," even acting together, could not prevent coral calcification rates on Meiji Reef from actually rising by about 11% over the past three centuries.
Focusing their attention on coral reefs spanning an 11° latitudinal range in the southeast Indian Ocean, Cooper et al. (2012) collected 27 long cores from massive Porites coral colonies at six locations covering a north-south distance of about 1000 km off the coast of Western Australia; and from these cores they developed 1900-2010 histories of "annual extension (linear distance between adjacent density minima, cm/year), skeletal density (g/cm3), and calcification rate (the product of skeletal density and annual extension, g/cm2/year)," based on gamma densitometry data.
In analyzing the cores, Cooper et al. report that at the Houtman Abrolhos Islands -- where a relatively large sea surface temperature (SST) increase had occurred (0.10°C/decade) -- calcification rates rose by 23.5%, and that at Coral Bay and Tantabiddi, SST increases of 0.8 and 0.6°C/decade were associated with 8.7 and 4.9% increases in decadal calcification rates, respectively, while smaller and non-significant positive trends in calcification rates were apparent at Clerke and Imperieuse Reefs, where the increase in SST was only 0.2°C/decade.
In discussing their observations, the three Australian researchers indicate that their latter non-significant findings are consistent with those of Helmle et al. (2011), who they say "found a similar non-correlation for the massive coral Montastraea faveolata in the Florida Keys between 1937 and 1996, when there was no significant SST warming." And they write - with respect to the large increases in calcification rates that they documented at the rapidly warming Houtman Abrolhos Islands - that "Lough and Barnes (2000) documented a similar positive correlation, suggesting that calcification rates may, at least initially, increase with global warming." Thus, they conclude that "the rate of change in the thermal environment of coral reefs is currently the primary driver of change in coral calcification rates," driving them ever higher as temperatures rise ever higher, and that "the large-scale phenomenon of ocean acidification is not currently limiting calcification on coral reefs uniformly at a global scale."
In prefacing their study, Hughes et al. (2012) write that "contemporary research on how climate change affects coral reefs has matured beyond the simplistic 'canary in the coal mine' concept to a more nuanced recognition that climate-related pressures such as bleaching (due to the loss of symbiotic zooxanthellae) and ocean acidification do not affect all species equally." And "in this context," as they continue, they indicate that "a critical issue for the future status of reefs will be their ability to maintain functional capacity in the face of the changes in species composition that are already underway due to multiple anthropogenic impacts."
To explore this subject in more detail, Hughes et al. applied a "rigorous quantitative approach to examine large-scale spatial variation in the species composition and abundance of corals on mid-shelf reefs along the length of Australia's Great Barrier Reef, a biogeographic region where species richness is high and relatively homogeneous." More specifically, they say they used "a hierarchical, nested sampling design to quantify scale-dependent patterns of coral abundances [for] five regions of the Great Barrier Reef [that they] sampled from north to south, each 250-500 km apart." Altogether, they thus identified and measured a total of 35,428 coral colonies on 33 reefs, categorizing each colony they encountered (including the majority of species that are too rare to analyze individually) into "ecologically relevant groups depending on their physiology, morphology and life history."
The seven scientists report that the diverse pool of species they examined along the latitudinal gradient of the Great Barrier Reef "can assemble in markedly different configurations across a wide range of contemporary environments." With respect to temperature, for example, they indicate that "the geographic ranges of 93% of the 416 coral species found on the Great Barrier Reef extend northwards toward the equator (e.g., to Papua New Guinea, the Solomon Islands, and/or the Indonesian archipelago)," while "46% are also found in colder conditions further to the south." As for ocean acidification, they state that "globally, ocean surface pH has decreased by 0.1 unit since 1750 due to the uptake of atmospheric CO2, with a smaller 0.06 decline recorded for the tropics," citing Kleypas et al. (2006). In contrast, however, they report that contemporary variation in pH among various reef habitats on the Great Barrier Reef, as well as differences among short-term replicate measurements, span a range of 0.39 unit, from 8.37 to 7.98, citing Gagliano et al. (2010). And they rightfully note that this short-term and habitat-scale variability literally swamps that of latitudinal trends.
In light of their illuminating real-world observations, Hughes et al. say that their results "all point to a surprisingly resilient response by some elements of coral assemblages to spatial and temporal shifts in climatic conditions." And, therefore, they contend that "the flexibility in community composition that we document along latitudinal environmental gradients indicates that climate change is likely to result in a re-assortment of coral reef taxa rather than wholesale loss of entire reef ecosystems." And this demonstrable state of marine life affairs pretty much deflates the catastrophic prognostications put forth by the IPCC with regard to the future of the planet's corals.
References
Bessat, F. and Buigues, D. 2001. Two centuries of variation in coral growth in a massive Porites colony from Moorea (French Polynesia): a response of ocean-atmosphere variability from south central Pacific. Palaeogeography, Palaeoclimatology, Palaeoecology 175: 381-392.
Browne, N.K. 2012. Spatial and temporal variations in coral growth on an inshore turbid reef subjected to multiple disturbances. Marine Environmental Research 77: 71-83.
Cantin, N.E., van Oppen, M.J.H., Willis, B.L., Mieog, J.C. and Negri, A.P. 2009. Juvenile corals can acquire more carbon from high-performance algal symbionts. Coral Reefs 28: 405-414.
Cooper, T.F., O'Leary, R.A. and Lough, J.M. 2012. Growth of Western Australian corals in the Anthropocene. Science 335: 593-596.
De'ath, G., Lough, J.M. and Fabricius, K.E. 2009. Declining coral calcification on the Great Barrier Reef. Science 323: 116-119.
Fabricius, K.E. 2005. Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Marine Pollution Bulletin 50: 125-146.
Fabricius, K.E., Langdon, C., Uthicke, S., Humphrey, C., Noonan, S., De'ath, G., Okazaki, R., Muehllehner, N., Glas, M.S. and Lough, J.M. 2011. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nature Climate Change 1: 165-169.
Gagliano, M., McCormick, M., Moore, J. and Depczynski, M. 2010. The basics of acidification: baseline variability of pH on Australian coral reefs. Marine Biology 157: 1849-1856.
Helmle, K.P., Dodge, R.E., Swart, P.K., Gledhill, D.K. and Eakin, C.M. 2011. Growth rates of Florida corals from 1937 to 1996 and their response to climate change. Nature Communications 2: 10.1038/ncomms1222.
Hoegh-Guldberg, O. 1999. Climate change, coral bleaching and the future of the world's coral reefs. Marine and Freshwater Research 50: 839-866.
Hoegh-Guldberg, O., Mumby, P.J., Hooten, A.J., Steneck, R.S., Greenfield, P., Gomez, E., Harvell, C.D., Sale, P.F., Edwards, A.J., Caldeira, K., Knowlton, N, Eakin, C.M., Iglesias-Prieto, R., Muthiga, N., Bradbury, R.H., Dubi, A. and Hatziolos, M.E. 2007. Coral reefs under rapid climate change and ocean acidification. Science 318: 1737-1742.
Hughes, T.P., Baird, A.H., Dinsdale, E.A., Moltschaniwskyj, N.A., Pratchett, M.S., Tanner, J.E. and Willis, B.L. 2012. Assembly rules of reef corals are flexible along a steep climatic gradient. Current Biology 22: 736-741.
Idso, S.B., Idso, C.D. and Idso, K.E. 2000. CO2, global warming and coral reefs: Prospects for the future. Technology 7S: 71-94.
Kleypas, J.A., Buddemeier, R.W., Archer, D., Gattuso, J.P., Langdon, C. and Opdyke, B.N. 1999. Geochemical consequences of increased atmospheric carbon dioxide on coral reefs. Science 284: 118-120.
Kleypas, J., Feeley, R.A., Fabry, V.J., Langdon, C., Sabine, C.L. and Robbins, L.L. 2006. Impacts of Ocean Acidification on Coral Reefs and Other Marine Calcifiers: A Guide for Future Research. Report of a workshop held April 18-20, 2005, St. Petersburg, Florida, USA, sponsored by NSF, NOAA and the U.S. Geological Survey.
Little, A.F., van Oppen, M.J.H. and Willis, B.L. 2004. Flexibility in algal endosymbioses shapes growth in reef corals. Science 304: 1492-1494.
Lough, J.M. and Barnes, D.J. 1997. Several centuries of variation in skeletal extension, density and calcification in massive Porites colonies from the Great Barrier Reef: A proxy for seawater temperature and a background of variability against which to identify unnatural change. Journal of Experimental and Marine Biology and Ecology 211: 29-67.
Lough, J.M. and Barnes, D.J. 2000. Environmental controls on growth of the massive coral Porites. Journal of Experimental Marine Biology and Ecology 245: 225-243.
McNeil, B.I., Matear, R.J. and Barnes, D.J. 2004. Coral reef calcification and climate change: The effect of ocean warming. Geophysical Research Letters 31: 10.1029/2004GL021541.
Pandolfi, J.M., Bradbury, R.H., Sala, E., Hughes, T.P., Bjorndal, K.A., Cooke, R.G., McArdle, D., McClenachan, L., Newman, M.J.H., Paredes, G., Warner, R.R. and Jackson, J.B.C. 2003. Global trajectories of the long-term decline of coral reef ecosystems. Science 301: 955-958.
Puotinen, M.L. 2004. Tropical Cyclone Impacts on Coral Reef Communities: Modeling the Disturbance Regime in the Great Barrier Reef Region, 1969-2003. Ph.D. Thesis. James Cook University, Townsville, Queensland, Australia.
Shi, Q., Yu, K.F., Chen, T.R., Zhang, H.L., Zhao, M.X. and Yan, H.Q. 2012. Two centuries-long records of skeletal calcification in massive Porites colonies from Meiji Reef in the southern South China Sea and its responses to atmospheric CO2 and seawater temperature. Science China Earth Sciences 55: 10.1007/s11430-011-4320-0.
Uthicke, S., Patel, F. and Ditchburn, R. 2012. Elevated land runoff after European settlement perturbs persistent foraminiferal assemblages on the Great Barrier Reef. Ecology 93: 111-121.
Wilkinson, C. 2004. Status of Coral Reefs of the World: 2004. Australian Institute of Marine Science Press, Townsville, Australia.
Last updated 14 October 2015