




In an early review of the direct effects of atmospheric CO2 enrichment on the growth of marine phytoplankton, Wolf-Gladrow et al. (1999) concluded that increasing atmospheric CO2 concentrations may lead to significant increases in phytoplanktonic growth rates, with a resultant tendency for the world's oceans to take up and store more atmospheric CO2 and thereby provide a means for the planet to counter the prospect of runaway global warming. In subsequent years, additional evidence for this conclusion has been obtained; and we here review some of the more recent pertinent findings.
Schippers et al. (2004a) developed "an integrated model of phytoplankton growth, air-water exchange and carbon chemistry to analyze the potential increase of phytoplankton productivity due to an atmospheric CO2 elevation," and to test the predictions of their model, they let the freshwater alga Chlamydomonas reinhardtii grow in 300-ml bottles filled with 150 ml of a nutrient-rich medium at enclosed atmospheric CO2 concentrations of 350 and 700 ppm, which they maintained at two air-water exchange rates as described by Shippers et al. (2004b), while they periodically measured the biovolume of the solutions by means of an electronic particle counter. Their results confirmed "the theoretical prediction that if algal effects on carbon chemistry are strong, increased phytoplankton productivity because of atmospheric CO2 elevation should become proportional to the increased atmospheric CO2," and these results suggested that although "strong algal effects (resulting in high pH levels) at which this occurs are rare under natural conditions," their model still predicted there would be (1) "a potential productivity increase of up to 40%, at observed pH levels for marine species with low affinity for HCO3-," and that (2) the effects on algal production in freshwater systems could potentially be larger, such that a doubling of atmospheric CO2 "may result in an increase of the productivity of more than 50%." Therefore, with respect to the ongoing rise in the air's CO2 content, Schippers et al. say their results suggest that "the aquatic carbon sink may increase more than expected," which would help to slow the rate-of-rise of the air's CO2 concentration and provide a greater food base for higher-trophic-level marine and freshwater organisms.
Contemporaneously, Riebesell (2004) presented some results of CO2 perturbation experiments conducted in a fjord south of Bergen, Norway, where nine 11-m3 enclosures moored to a floating raft were aerated in triplicate with CO2-depleted, normal, and CO2-enriched air to achieve CO2 levels of 190, 370 and 710 ppm, simulating glacial, present day, and predicted conditions for the end of the century, respectively. In the course of the study, he reported that a bloom consisting of a mixed phytoplankton community developed; and, in his words, "significantly higher net community production was observed under elevated CO2 levels during the build-up of the bloom." In addition, he stated that "CO2-related differences in primary production continued after nutrient exhaustion, leading to higher production of transparent exopolymer particles under high CO2 conditions," something that had also been observed by Engel (2002) in a natural plankton assemblage. These particles, according to Riebesell, "accelerate particle aggregation and thereby enhance vertical particle flux," which he says may "provide an efficient pathway to channel dissolved and colloidal organic matter into the particulate pool," which would also tend to increase the "aquatic carbon sink" discussed by Schippers et al.
Three years later, Riebesell et al. (2007) reported the findings of another experiment that was conducted at the same location, where they maintained nine cylindrical mesocosms that extended from the water surface to a depth of 9-10 meters in equilibrium with air of either ambient CO2 concentration (350 ppm), doubled CO2 (700 ppm) or tripled CO2 (1050 ppm), while they measured several phytoplanktonic physiological parameters. This study revealed, as they describe it, that "net community carbon consumption under increased CO2 exceeded present rates by 27% (2 x CO2) and 39% (3 x CO2)," and they say that continuous oxygen measurements in the mesocosms indicated "enhanced net photosynthesis to be the source of the observed CO2 effect."
Commenting on their findings, the eleven researchers said they "underscore the importance of biologically driven feedbacks in the ocean to global change," while Arrigo (2007) stated in a News & Views discussion of Riebesell et al.'s paper that "increased CO2 has been shown to enhance fixation of free nitrogen, thereby relaxing nutrient limitation by nitrogen availability and increasing CO2 uptake (Barcelos e Ramos et al., 2007)," further noting that such non-steady-state biological feedbacks are currently not included in models of global climate, while indicating that the phytoplanktonic growth-promoting effect of CO2 observed by Riebesell et al. has probably been responsible for limiting the rise in atmospheric CO2 experienced since the dawn of the Industrial Revolution to approximately 90% of what it likely would have been in its absence.
The following year, Tortell et al. (2008) measured CO2 uptake of in situ phytoplankton assemblages collected at 35 stations in the Ross Sea polynya during Austral spring and summer, together with 14C uptake for a subset of 11 station samples, while they conducted CO2 manipulation experiments with phytoplankton collected at three Ross Sea locations via shipboard incubations using a semi-continuous batch-culture technique. This work revealed that for springtime phytoplankton assemblages there was a statistically significant increase in 14C fixation between 100 and 380 ppm CO2, but no further effects observed at 800 ppm CO2. In the case of summertime phytoplankton assemblages, on the other hand, the CO2-induced increase in both relative growth rate and primary productivity continued all the way out to the highest CO2 concentration investigated, i.e., 800 ppm, while additionally promoting "a shift towards larger chain-forming species." And in writing of these observations, Tortell et al. state that "potential CO2-dependent productivity increases and algal species shifts could thus act to increase the efficiency of the biological pump, enhancing Southern Ocean CO2 uptake and contributing to a negative feedback on increased atmospheric CO2."
Around the same time, Wu et al. (2008) reviewed what was known about the effects of atmospheric CO2 enrichment on both micro- and macro-algae living in the world's oceans, finding that air enriched with CO2 up to several times the present atmospheric level had been shown "to enhance photosynthesis and growth of both phytoplanktonic and macro-species that have less capacity of CCMs [CO2-concentrating mechanisms]," while adding that "even for species that operate active CCMs and those whose photosynthesis is not limited by CO2 in seawater, increased CO2 levels can down-regulate their CCMs and therefore enhance their growth under light-limiting conditions," because "at higher CO2 levels, less light energy is required to drive CCM." In addition, they reported that elevated CO2 levels had been found to enhance the activity of nitrogen reductase in several marine plants, and that this phenomenon "would support enhanced growth rate by providing adequate nitrogen required for their metabolism under the high CO2 level." Last of all, they noted that "altered physiological performances under high-CO2 conditions may cause genetic alteration in view of adaptation over long time scales," and that "marine algae may adapt to a high CO2 oceanic environment so that evolved communities in [the] future are likely to be genetically different from contemporary communities."
Also publishing in the same year were Vogt et al. (2008), who wrote that "oceanic DMS [dimethylsulphide] production is a result of complex interactions within the marine food-web," and that "ocean acidification may affect DMS concentrations and fluxes by altering one or more of the various pathways or impacting some of the species involved," with the reason for concern being the fact that the particulate atmospheric oxidation products of DMS can act as cloud condensation nuclei and lead to the creation of more numerous and more reflective clouds that can cool the planet by reflecting more incoming solar radiation back to space, which would tend to mute the greenhouse effect of rising atmospheric CO2 concentrations and keep the planet from getting too warm. Therefore, they studied the effects of atmospheric CO2 enrichment on various marine microorganisms and DMS production in nine marine mesocosms maintained within 2-meter-diameter polyethylene bags submerged to a depth of ten meters in a fjord adjacent to the Large-Scale Facilities of the Biological Station of the University of Bergen in Espegrend, Norway, where three of the mesocosms were maintained at ambient levels of CO2 (~375 ppm), three were maintained at levels expected to prevail at the end of the current century (760 ppm or 2x CO2), and three were maintained at levels predicted for the middle of the next century (1150 ppm or 3x CO2), while measurements of numerous ecosystem parameters were made over a period of 24 days.
In reporting their results, Vogt et al. say they detected no significant phytoplankton species shifts among treatments, and that "the ecosystem composition, bacterial and phytoplankton abundances and productivity, grazing rates and total grazer abundance and reproduction were not significantly affected by CO2 induced effects," citing in further support of this statement the similar findings of Riebesell et al. (2007), Riebesell et al. (2008), Egge et al. (2007), Paulino et al. (2007), Larsen et al. (2007), Suffrian et al. (2008) and Carotenuto et al. (2007). In addition, they say that "while DMS stayed elevated in the treatments with elevated CO2," they observed "a steep decline in DMS concentration in the treatment with low CO2," i.e., the ambient treatment. Consequently, the eight researchers demonstrated that the marine biological system they studied "was surprisingly resilient to abrupt and large pH changes," which is just the opposite of what the world's climate alarmists characteristically predict about CO2-induced "ocean acidification." And that may be why Vogt et al. described the ecosystem they studied as being "surprisingly resilient."
Working at the same site, after a one-time addition of nutrients that were used to initiate a phytoplankton bloom on the day prior to the start of their 24-day study, Egge et al. (2009) measured primary production in the nine mesocosms at two-day intervals, along with oxygen production and consumption, the presence of transparent exopolymer particles, and the composition of the phytoplanktonic community. This work revealed that "in the second half of the experiment there was a tendency of higher production at elevated CO2 levels," which was "visible from ca. day 10 in the cumulative production, with a significant difference between 3x and 1x CO2 from day 20 onward," as shown in the figure below.
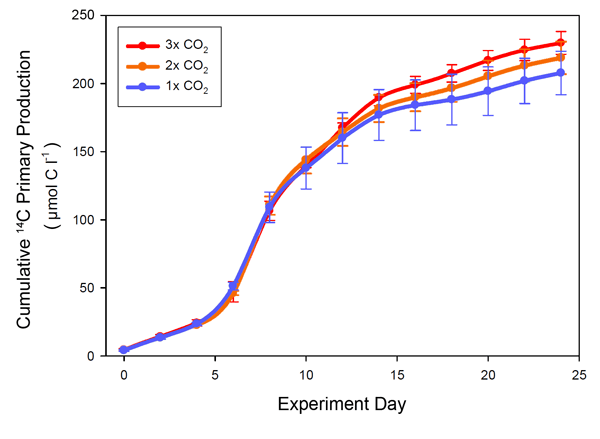
Cumulative primary production from the start of the experiment. Adapted from Egge et al. (2009).
Egge et al. thus concluded that their results "demonstrate a small, but statistically significant effect of elevated CO2 on daily primary production," which they describe as being "consistent with the over-consumption of dissolved inorganic carbon at elevated CO2 reported by Riebesell et al. (2007) and Bellerby et al. (2008)." Consequently, their observations once again suggest that the planet's rising atmospheric CO2 concentration may well stimulate oceanic primary production and thereby enable the sustaining of a greater population of higher-trophic-level marine organisms, many of which could ultimately end up on our dinner tables.
References
Barcelos e Ramos, J., Biswas, H., Schulz, K.G., LaRoche, J. and Riebesell, U. 2007. Effect of rising atmospheric carbon dioxide on the marine nitrogen fixer Trichodesmium. Global Biogeochemical Cycles 21: 10.1029/2006GB002898.
Bellerby, R.G.J., Schulz, K.G., Riebesell, U., Neil, C., Nondal, G., Johannessen, T. and Brown, K.R. 2008. Marine ecosystem community carbon and nutrient uptake stoichiometry under varying ocean acidification during the PeECE III experiment. Biogeosciences 5: 1517-1527.
Carotenuto, Y., Putzeys, S., Simonelli, P., Paulino, A., Meyerhofer, M., Suffrian, K., Antia, A. and Nejstgaard, J.C. 2007. Copepod feeding and reproduction in relation to phytoplankton development during the PeECE III mesocosm experiment. Biogeosciences Discussions 4: 3913-3936.
Egge, J., Thingstad, F., Engel, A., Bellerby, R.G.J. and Riebesell, U. 2007. Primary production at elevated nutrient and pCO2 levels. Biogeosciences Discussions 4: 4385-4410.
Egge, J.K, Thingstad, T.F., Larsen, A., Engel, A., Wohlers, J., Bellerby, R.G.J. and Riebesell, U. 2009. Primary production during nutrient-induced blooms at elevated CO2 concentrations. Biogeosciences 6: 877-885.
Engel, A. 2002. Direct relationship between CO2 uptake and transparent exopolymer particles production in natural phytoplankton. Journal of Plankton Research 24: 49-53.
Larsen, J.B., Larsen, A., Thyrhaug, R., Bratbak, G. and Sandaa R.-A. 2007. Marine viral populations detected during a nutrient induced phytoplankton bloom at elevated pCO2 levels. Biogeosciences Discussions 4: 3961-3985.
Paulino, A.I., Egge, J.K. and Larsen, A. 2007. Effects of increased atmospheric CO2 on small and intermediate sized osmotrophs during a nutrient induced phytoplankton bloom. Biogeosciences Discussions 4: 4173-4195.
Riebesell, U. 2004. Effects of CO2 enrichment on marine phytoplankton. Journal of Oceanography 60: 719-729.
Riebesell, U., Bellerby, R.G.J., Grossart, H.-P. and Thingstad, F. 2008. Mesocosm CO2 perturbation studies: from organism to community level. Biogeosciences Discussions 5: 641-659.
Riebesell, U., Schulz, K.G., Bellerby, R.G.J., Botros, M., Fritsche, P., Meyerhofer, M., Neill, C., Nondal, G., Oschlies, A., Wohlers, J. and Zollner, E. 2007. Enhanced biological carbon consumption in a high CO2 ocean. Nature 450: 545-548.
Schippers, P., Lurling, M. and Scheffer, M. 2004a. Increase of atmospheric CO2 promotes phytoplankton productivity. Ecology Letters 7: 446-451.
Schippers, P., Vermaat, J.E., de Klein, J. and Mooij, W.M. 2004. The effect of atmospheric carbon dioxide elevation on plant growth in freshwater ecosystems. Ecosystems 7: 63-74.
Suffrian, K., Simonelli, P., Nejstgaard, J.C., Putzeys, S., Carotenuto, Y. and Antia, A.N. 2008. Microzooplankton grazing and phytoplankton growth in marine mesocosms with increased CO2 levels. Biogeosciences Discussions 5: 411-433.
Tortell, P.D., Payne, C.D., Li, Y., Trimborn, S., Rost, B., Smith, W.O., Riesselman, C., Dunbar, R.B., Sedwick, P. and DiTullio, G.R. 2008. CO2 sensitivity of Southern Ocean phytoplankton. Geophysical Research Letters 35: 10.1029/2007GL032583.
Vogt, M., Steinke, M., Turner, S., Paulino, A., Meyerhofer, M., Riebesell, U., LeQuere, C. and Liss, P. 2008. Dynamics of dimethylsulphoniopropionate and dimethylsulphide under different CO2 concentrations during a mesocosm experiment. Biogeosciences 5: 407-419.
Wolf-Gladrow, D.A., Riebesell, U., Burkhardt, S. and Bijma, J. 1999. Direct effects of CO2 concentration on growth and isotopic composition of marine plankton. Tellus 51B: 461-476.
Wu, H.-Y., Zou, D.-H. and Gao, K.-S. 2008. Impacts of increased atmospheric CO2 concentration on photosynthesis and growth of micro- and macro-algae. Science in China Series C: Life Sciences 51: 1144-1150.
Last updated 28 July 2010