




Foraminifera are amoeboid protists with reticulating pseudopods, i.e., fine strands of cytoplasm that subdivide into branches that merge to form a dynamic network. They are typically less than one mm in size (but can be much larger), and they produce an elaborate calcium carbonate shell called a test, which may have one or more chambers. As for their impact on the undersea marine environment, these widespread calcifying protozoa, acording to Schiebel (2002), are responsible for 32-80% of the global deep-ocean flux of calcite. Therefore, it is important to determine the degree to which various forams -- as they are often called -- may or may not be harmed by likely future increases in what has come to be known as ocean acidification.
Working with specimens of Orbulina universa collected by scuba divers off the coast of Catalina Island, California (USA), plus Globigerinoides sacculifer obtained near Puerto Rico (USA), Lombard et al. (2010) cultured the two forams under high and low irradiances (335 and 35 umol photons m-2 s-1, respectively) in filtered seawater that had its pH and carbonate ion concentration (CO32-) manipulated by adding NaOH or HCl to it, while they measured the forams' initial and final size, survival time (days from collection to gametogenesis), and final shell weight, but only for individuals that underwent gametogenesis and grew at least one chamber. And in doing so, they determined that under the IS92a "business as usual" scenario as defined by the Intergovernmental Panel on Climate Change and projected for the year 2100, the calcification rates of G. sacculifer and O. universa "could decline by 6-13% compared to recent rates."
Clearly, 6-13% is not a large decline. In addition, the four researchers say that "the future increase in temperature [predicted by the IPCC] could increase the production of calcite by foraminifera, counteracting the negative impact of ocean acidification," which could well reduce the expected decline in calcification to next to nothing. And the results of the analysis of Tans (2009), which are included in the figure below, suggest that (1) the true decline in oceanic pH by the year 2100 is more likely to be only about half of that projected by the IPCC, and that (2) this drop will begin to be ameliorated shortly after 2100, gradually returning oceanic pH to present-day values beyond AD 2500. And in light of these several considerations, it could well be that climate-alarmist "concerns" about CO2-induced ocean acidification are but a tempest in a teapot and/or much ado about next to nothing -- in the cases of these two forams, at least.
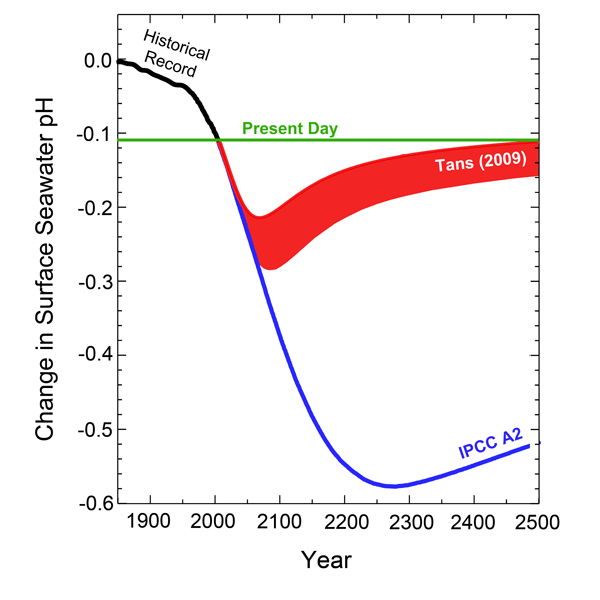
Figure 1. The change in surface seawater pH vs. time, as observed for the prior century and a half and as calculated by the IPCC and Tans (2009) for the next five centuries.
In another foraminifera study, Kuroyanagi et al. (2009) cultured asexually-produced individuals of Marginopora kudakajimensis -- a large calcifying microorganism that contributes to both organic and inorganic carbon production in coral reefs -- under carefully controlled laboratory conditions for a period of 71 days in glass jars containing approximately 110 ml of filtered natural seawater (control pH of about 8.2) and two less basic pH conditions of about 7.9 and 7.7, which lower values were created by additions of 0.1 N HCl. This work revealed that in declining from the control pH of 8.2 to a pH of 7.9, the mean maximum shell diameter of the large foraminifer actually rose by 8.6%, while its mean shell weight rose by a much smaller and insignificant 0.7%. As the seawater's pH declined all the way to 7.7, however, the organism's mean maximum shell diameter fell by 12.1%, while its mean shell weight fell by 49.3%.
Based on these observations, Kuroyanagi et al. concluded that if oceanic pH remains within the range of 8.2 to 7.9, "large foraminifers should be able to maintain present calcification rates," but they also speculated that any further drop in pH could lead to reduced rates of calcification. We hasten to add in this regard, however, that although the IPCC's A2 scenario predicts a maximum pH decline of approximately 0.5 pH units by about AD 2270, the more recent analysis of Tans (2009) suggests a maximum pH drop of only about 0.14 unit by about AD 2090, after which pH begins to rise asymptotically to return to its current value after a few additional centuries. And this latter projection suggests that oceanic pH will never even come close to creating a major decline in M. kudakajimensis calcification rate.
Contemporaneously, Bernhard et al. (2009) grew the marine foraminiferal protist Allogromia laticollaris -- which they describe as "a ubiquitous protistan constituent of marine microbial systems" and "an important link in the marine food web" -- in a mixture of 32%0 seawater and Alga-Gro seawater medium in 20-ml glass culture tubes, while examining its response to a number of super-high atmospheric CO2 concentrations to which the tubes were exposed: 15,000, 30,000, 60,000, 90,000 and 200,000 ppm, which values are to be compared to the study's atmospheric control concentration of 375 ppm CO2. This work revealed, in their words, that the protist they studied "is able to survive 10-14-day exposure to elevated CO2 as high as 200,000 ppm." In fact, they say that "both ATP [Adenosine Triphosphate, an indicator of cellular energy] data and microscopic examination indicate that considerable populations of A. laticollaris survived exposure to all experimental treatments of elevated CO2, even both replicates of the 200,000-ppm CO2 experiments [italics added]." And they found that "at least three specimens reproduced during exposure to either 90,000 ppm or 200,000 ppm CO2," while "such reproduction was observed only once in an atmospheric [375-ppm CO2] treatment."
In light of these observations, the four researchers state that "A. laticollaris is an appropriate species to predict the response of shallow-water thecate foraminifera to predicted increases in atmospheric CO2, given its isolation [i.e., acquisition] from a shallow-water semi-tropical setting." Hence, they go on to say their results suggest that "at least some foraminiferal species will tolerate CO2 values that are one to two orders of magnitude higher than those predicted for the next few centuries [italics added]." And, last of all, they say that A. laticollaris will also tolerate CO2 values that are one to two orders of magnitude greater than those predicted to occur for the "extreme case" of burning all fossil fuels in the crust of the earth, which suggests that A. laticollaris is, indeed, one tough protist!
But aren't all forams amazingly resilient creatures?
In broaching this question, we briefly review the findings of Cannariato et al. (1999), who investigated the character, magnitude and speed of biotic responses of benthic foraminifera to millennial-scale climatic oscillations, using data obtained from an ocean sediment core in the Northeast Pacific that extended back in time some 60,000 years. This work revealed a number of rapid climatic switches throughout the course of the 60,000-year record, representing periods of what the three researchers called "extreme environmental variability." However, they report that no extinctions were ever observed and that the benthic ecosystems "appear to be both resilient and robust in response to rapid and often extreme environmental conditions," stating that faunal turnovers occurred within decades throughout the record "without extinction or speciation." As a result, they concluded that "broad segments of the biosphere are well adapted to rapid climate change," which further suggests that broad segments of the biosphere may be equally well adapted to the environmental change of ocean acidification, which climate alarmists associate with the ongoing rise in the air's CO2 content, but which Tans (2009) has shown to be much less extreme than they originally hypothesized.
References
Bernhard, J.M., Mollo-Christensen, E., Eisenkolb, N. and Starczak, V.R. 2009. Tolerance of allogromiid Foraminifera to severely elevated carbon dioxide concentrations: Implications to future ecosystem functioning and paleoceanographic interpretations. Global and Planetary Change 65: 107-114.
Cannariato, K.G., Kennett, J.P. and Behl, R.J. 1999. Biotic response to late Quaternary rapid climate switches in Santa Barbara Basin: Ecological and evolutionary implications. Geology 27: 63-66.
Kuroyanagi, A., Kawahata, H., Suzuki, A., Fujita, K. and Irie, T. 2009. Impacts of ocean acidification on large benthic foraminifers: Results from laboratory experiments. Marine Micropaleontology 73: 190-195.
Lombard, F., da Rocha, R.E., Bijma, J. and Gattuso, J.-P. 2010. Effect of carbonate ion concentration and irradiance on calcification in planktonic foraminifera. Biogeosciences 7: 247-255.
Schiebel, R. 2002. Planktic foraminiferal sedimentation and the marine calcite budget. Global Biogeochemical Cycles 16: 1010.1029/2001GB001459.
Tans, P. 2009. An accounting of the observed increase in oceanic and atmospheric CO2 and an outlook for the future. Oceanography 22: 26-35.
Last updated 14 July 2010