




Among the many climate-alarmist fears of CO2-induced global warming is the concern that the productivity of the biosphere will decline if global temperatures rise to the extent predicted by computer models. Yet, for many alarmists, the future is the present. Since 1980, for example, the Earth has weathered three of the warmest decades in the instrumental temperature record, a handful of intense and persistent El Niño events, large-scale deforestation, "unprecedented" forest fires, and the eruption of several volcanoes. Concurrently, the air's CO2 content increased by 16%, while human population grew by 55%. So just how bad is the biosphere suffering in response to these much-feared events? Or, is it even suffering at all?
In the following summary, we explore the findings of several research groups that have set out to answer such questions, specifically focusing on the time period of the past few decades for the Earth as a whole. For according to the alarmists, if there ever was a time in which the biosphere should be showing signs of distress, it most certainly would be the recent past.
We begin with the study of Reichenau and Esser (2003), who investigated the individual influences of ENSO, volcanic eruptions and the North Atlantic Oscillation (NAO) on the 1958-1994 time series of anomalous CO2 fluxes with the help of the High Resolution Biosphere Model of Esser et al. (1994) and real-world CO2 measurements. Although the two researchers could conclude nothing about the NAO, other than to say "the influence of the NAO remain[s] unclear," they were able to determine that periods of anomalous rising atmospheric CO2 concentrations coincided with El Niño periods, while periods of anomalous declining atmospheric CO2 concentrations coincided with periods of significant volcanism.
Enlarging on these observations, they say that "the globally averaged effect of [the El Niño] circulation pattern on the terrestrial biosphere is a net release of carbon," in agreement with the results of earlier investigations of the subject (Bacastow, 1976; Bacastow et al., 1980), which in turn "confirms earlier findings that the terrestrial biosphere is mainly responsible for atmospheric CO2 variations on the ENSO timescale (Keeling et al., 1995; Lee et al., 1998; Feely et al., 1999; Gerard et al., 1999; Rayner and Law, 1999; Battle et al., 2000; Bousquet et al., 2000; Houghton, 2000; Knorr, 2000, Le Quere et al., 2000; Langenfelds et al., 2002)." At the other end of the spectrum, Reichenau and Esser report that "volcanic eruptions with considerable aerosol production may create disturbances of the (biospheric) carbon cycle by increasing the photosynthetic carbon uptake due to the enhanced diffuse fraction of the incoming [solar] radiation," which accords with the findings of Roderick et al. (2001), Cohan et al. (2002), Law et al. (2002) and Gu et al. (2002, 2003).
The many published studies of anomalous CO2 fluxes between Earth's surface and its atmosphere clearly indicate that warm El Niño conditions tend to reduce biospheric productivity, while cool volcanic conditions tend to enhance biospheric productivity. Does this observation imply that "cool is good" and "warm is bad" for the planet's plants?
No, it does not. For one thing, the productivity-enhancing effect of volcanic eruptions arises not from their cooling influence, but from their increasing the amount of diffuse solar radiation received at the Earth's surface, which allows for an enhanced penetration of solar radiation deeper into plant canopies, which reduces within-canopy shade and boosts rates of canopy net photosynthesis. As for the productivity-reducing effect of El Niños, it could well be more a consequence of changes in global precipitation patterns than of the direct effect of an increase in temperature. It is known from the work of Indermuhle et al. (1999), for example, that the pattern of biospheric productivity over the last 7,000 years of the Holocene was essentially that of a slow monotonic decline from the peak growth conditions of the interglacial's Climatic Optimum, which productivity decline, in their words, was "due to a change from the warmer and wetter mid-Holocene climate to colder and drier conditions."
Additional evidence of the benefits of long-term warming on plants comes from the study of Lin et al. (2010), who conducted a meta-analysis of data they obtained from 127 individual studies and found that for the totality of terrestrial plants included in their analysis, "warming significantly increased biomass by 12.3%," while noting there was a "significantly greater stimulation of woody (+26.7%) than herbaceous species (+5.2%)." They also found that the warming effects on plant biomass production "did not change with mean annual precipitation or experimental duration," and that "other treatments, including CO2 enrichment, nitrogen addition, drought and water addition, did not alter warming responses of plant biomass." As a result, the Chinese researchers conclude, in their words, that "results in this and previous meta-analyses (Arft et al., 1999; Rustad et al., 2001; Dormann and Woodin, 2001; Walker et al., 2006) have revealed that warming generally increases terrestrial plant biomass, indicating enhanced terrestrial carbon uptake via plant growth and net primary productivity."
Viewed from these perspectives, it can be appreciated that long-term global warming tends to substantially increase biospheric productivity. Short-term deviations from this basic relationship that are evident in anomalous variations of CO2 fluxes between the Earth's surface and atmosphere are just that: short-lived and anomalous.
Also examining carbon fluxes was Langenfelds et al. (1999), who analyzed O2/N2 measurements of the contents of a suite of tanks filled with background air collected at Cape Grim, Tasmania between April 1978 and January 1997. The rates of carbon storage in the world's oceans and the terrestrial biosphere they derived from these data indicated the terrestrial biosphere was essentially in balance with respect to surface fluxes of carbon throughout this 19-year interval. However, it is known from other studies that tropical deforestation produced a huge net loss of carbon during each of those years. As a result, Langenfelds et al. necessarily acknowledged the existence of a terrestrial carbon sink of like magnitude. This "compensating growth of the [terrestrial] biosphere," as they describe it, was suggested by them to be due to "reforestation, higher rates of net production in response to climatic trends, fertilisation by elevated levels of atmospheric CO2 or nitrogen deposition or a combination of these factors." Thus, although the physician is often accused of failing to heal himself, the biosphere appears to be doing just fine in this regard, yearly re-sequestering all of the carbon that man takes out of it.
With respect to the "fertilization of elevated levels of atmospheric CO2," in a broad review of the scientific literature, Idso and Idso (2011) described a number of biological consequences of this ongoing phenomena, a byproduct of the continued combustion of fossil fuels. The best known of these important impacts is probably CO2's aerial fertilization effect, which works its wonders on plants that utilize all three of the major biochemical pathways of photosynthesis (C3, C4 and CAM). In the case of herbaceous plants, this phenomenon typically boosts their productivities by about a third in response to a 300 ppm increase in the air's CO2 content, while it enhances the growth of woody plants by 50% or more.
Next comes plant water use efficiency, which may be defined as the amount of organic matter produced per unit of water transpired to the atmosphere. This parameter is directly enhanced by the aerial fertilization effect of atmospheric CO2 enrichment, as well as by its anti-transpirant effect, which is produced by CO2-induced decreases in the number density and degree of openness of leaf stomatal apertures that occur at higher atmospheric CO2 concentrations. Here, too, CO2-induced percentage increases as large as, or even larger than, those exhibited by plant productivity are commonplace.
One of the important ramifications of this CO2-induced increase in plant water use efficiency is the fact that it enables plants to grow and reproduce in areas that were previously too dry for them. With consequent increases in ground cover in these regions, the adverse effects of wind- and water-induced soil erosion are also reduced. Hence, there is a tendency for desertification to be reversed and for vast tracts of previously unproductive land to become supportive of more abundant animal life, both above- and below-ground, in what could appropriately be called a "greening of the Earth."
In addition to helping vegetation overcome the stress of limited water supplies, elevated levels of atmospheric CO2 help plants to better cope with other environmental stresses, such as low soil fertility, low light intensity, high soil and water salinity, high air temperature, various oxidative stresses and the stress of herbivory. When confronted with the specter of global warming, for example, many experiments have revealed that concomitant enrichment of the air with CO2 tends to increase the temperature at which plants function at their optimum, often making them even better suited to the warmer environment than they were to the cooler environment to which they were originally adapted. Under the most stressful of such conditions, in fact, extra CO2 sometimes is the deciding factor in determining whether a plant lives or dies.
These benefits of atmospheric CO2 enrichment apply to both agricultural and natural ecosystems; and as Wittwer (1995) has noted, "the rising level of atmospheric CO2 could be the one global natural resource that is progressively increasing food production and total biological output in a world of otherwise diminishing natural resources of land, water, energy, minerals, and fertilizer." This phenomenon is thus a means, he says, "of inadvertently increasing the productivity of farming systems and other photosynthetically active ecosystems," and that "the effects know no boundaries and both developing and developed countries are, and will be, sharing equally."
In light of these several observations, plus the fact that the air's CO2 content has risen substantially over the past two centuries - especially since 1950 - one would expect to see some evidence of the "greening of the Earth" (Idso, 1986) that they imply. And more and more researchers like Langenfelds et al. are acknowledging these stimulatory effects on Earth's biosphere.
In another approach to the subject, Idso (1995) laid out the evidence for a worldwide increase in the growth rates of Earth's forests that has been coeval with the progression of the Industrial Revolution and the rising CO2 content of the atmosphere. The development of this concept began with the study of LaMarche et al. (1984), who analyzed annual growth rings of two species of pine tree growing near the timberline in California, Colorado, Nevada and New Mexico (USA), and who thereby discovered large increases in growth rate between 1859 and 1983, which rates exceeded what might have been expected from climatic trends but were consistent with the global trend of atmospheric CO2. The developmental journey then continued with a study of ring-width measurements of Douglas fir trees in British Columbia, Canada, that also revealed a marked increase in growth in the trees' latter decades (Parker et al., 1987), leading the principal investigator of the project to state that "environmental influences other than increased CO2 have not been found that would explain this [phenomenon]." West (1988) reported much the same thing with respect to long-leaf pines in Georgia, i.e., that their annual growth increments had begun to rise at an unusual rate about 1920, increasing by approximately 30% by the mid-1980s; and he too stated that "the increased growth cannot be explained by trends in precipitation, temperature, or Palmer Drought Severity Index," leaving the rising CO2 content of the atmosphere as the likely cause of the increase in productivity.
Contemporaneously, stands of Scots pines in northern Finland were found to have experienced growth increases ranging from 15 to 43% between 1950 and 1983 (Hari et al., 1984; Hari and Arovaara, 1988). As to the cause of this phenomenon, the researchers stated that "CO2 seems to be the only environmental factor that has been changing systematically during this century in the remote area under study," and it was thus to this factor that they looked for an explanation of their observations.
The next major development in the continuing saga was the finding of Graybill and Idso (1993) that very long ring-width chronologies (some stretching back nearly 1800 years) of high-altitude long-lived bristlecone, foxtail and limber pine trees in Arizona, California, Colorado and Nevada (USA) all developed an unprecedented upward growth trend somewhere in the 1850s that continued as far towards the present as the records extended. In this case, too, like the ones that preceded it, comparisons of the chronologies with temperature and precipitation records ruled out the possibility that either of these climatic variables played a significant role in enhancing the trees' growth rates, strongly implicating the historical rise in the air's CO2 content as the factor responsible for their ever-increasing productivity over the prior century and a half.
Perhaps the most striking evidence of all for the significant 20th-century growth enhancement of earth's forests by the historical increase in the air's CO2 concentration was provided by the study of Phillips and Gentry (1994). Noting that turnover rates of mature tropical forests correlate well with measures of net productivity (Weaver and Murphy, 1990), the two scientists assessed the turnover rates of 40 tropical forests from around the world in order to test the hypothesis that global forest productivity was increasing in situ. In doing so, they found that the turnover rates of these highly productive forests had indeed been rising ever higher since at least 1960, with an apparent pan-tropical acceleration since 1980. In discussing what might be causing this phenomenon, they stated that "the accelerating increase in turnover coincides with an accelerating buildup of CO2," and as Pimm and Sugden (1994) stated in a companion article, it was "the consistency and simultaneity of the changes on several continents that lead Phillips and Gentry to their conclusion that enhanced productivity induced by increased CO2 is the most plausible candidate for the cause of the increased turnover."
Four years later, a group of eleven researchers headed by Phillips (Phillips et al., 1998) reported another impressive finding. Working with data on tree basal area (a surrogate for tropical forest biomass) for the period 1958-1996, which they obtained from several hundred plots of mature tropical trees scattered about the world, they found that average forest biomass for the tropics as a whole had increased substantially. In fact, they calculated that the increase amounted to approximately 40% of the missing terrestrial carbon sink of the entire globe. Hence, they suggested that "intact forests may be helping to buffer the rate of increase in atmospheric CO2, thereby reducing the impacts of global climate change," as Idso (1991a,b) had earlier suggested, and they identified the aerial fertilization effect of the ongoing rise in the air's CO2 content as one of the factors responsible for this phenomenon. Other contemporary studies also supported their findings (Grace et al., 1995; Malhi et al., 1998), verifying the fact that neotropical forests were indeed accumulating ever more carbon; and Phillips et al. (2002) continued to state that this phenomenon was occurring "possibly in response to the increasing atmospheric concentrations of carbon dioxide (Prentice et al., 2001; Malhi and Grace, 2000)."
As time progressed, however, it became less and less popular (i.e., ever more politically incorrect) to report positive biological consequences of the ongoing rise in the air's CO2 concentration; and the conclusions of Phillips and company began to be repeatedly challenged (Sheil, 1995; Sheil and May, 1996; Condit, 1997; Clark, 2002; Clark et al., 2003). In response to those challenges, Phillips, joined by 17 other researchers (Lewis et al., 2005), including one who had earlier criticized his and his colleagues' conclusions, published a new analysis that vindicated Phillips et al.'s earlier thoughts on the subject.
One of the primary concerns of the critics of Phillips et al.'s work was that their meta-analyses included sites with a wide range of tree census intervals (2-38 years), which they claimed could be confounding or "perhaps even driving conclusions from comparative studies," as Lewis et al. (2005) describe it. However, in Lewis et al.'s detailed study of this potential problem, which they concluded was indeed real, they found that re-analysis of Phillips et al.'s published results "shows that the pan-tropical increase in stem turnover rates over the late 20th century cannot be attributed to combining data with differing census intervals." Or as they state more obtusely in another place, "the conclusion that turnover rates have increased in tropical forests over the late 20th century is robust to the charge that this is an artifact due to the combination of data that vary in census interval (cf. Sheil, 1995)."
Lewis et al. (2005) additionally noted that "Sheil's (1995) original critique of the evidence for increasing turnover over the late 20th century also suggests that the apparent increase could be explained by a single event, the 1982-83 El Niño Southern Oscillation (ENSO), as many of the recent data spanned this event." However, as they continued, "recent analyses from Amazonia have shown that growth, recruitment and mortality rates have simultaneously increased within the same plots over the 1980s and 1990s, as has net above-ground biomass, both in areas largely unaffected, and in those strongly affected, by ENSO events (Baker et al., 2004; Lewis et al., 2004; Phillips et al., 2004)."
In a review of these several global forest studies, as well as many others (which led to their citing 186 scientific journal articles), Boisvenue and Running (2006) examined reams of "documented evidence of the impacts of climate change trends on forest productivity since the middle of the 20th century." In doing so, they found that "globally, based on both satellite and ground-based data, climatic changes seemed to have a generally positive impact on forest productivity when water was not limiting," which was most of the time, because they report that "less than 7% of forests are in strongly water-limited systems." Hence, and in spite of what climate alarmists routinely describe as unprecedented increases in the "twin evils" of rising atmospheric CO2 concentrations and air temperatures, there has in fact been what Boisvenue and Running call a significant "greening of the biosphere," and the world's forests in particular.
Three years later in another review of the scientific literature pertaining to this important issue of global biospheric productivity, Lewis et al. (2009) evaluated tropical forest inventory data, plant physiology experiments, ecosystem flux observations, Earth observations, atmospheric measurements and dynamic global vegetation models. In doing so, the five researchers report that both theory and experiments suggest that over the past several decades "plant photosynthesis should have increased in response to increasing CO2 concentrations, causing increased plant growth and forest biomass." And in this regard they find that "long-term plot data collectively indicate an increase in carbon storage, as well as significant increases in tree growth, mortality, recruitment, and forest dynamism." They also say that satellite measurements "indicate increases in productivity and forest dynamism," and that five Dynamic Global Vegetation Models, incorporating plant physiology, competition, and dynamics, all predict increasing gross primary productivity, net primary productivity, and carbon storage when forced using late-twentieth century climate and atmospheric CO2 concentration data." In addition, they state that "the predicted increases in carbon storage via the differing methods are all of similar magnitude (0.2% to 0.5% per year)."
Another intriguing indication that these CO2- and temperature-induced biological benefits are indeed in process of occurring in the real world of nature comes from the decline in the air's diurnal temperature range (DTR) that is evident in many parts of the world (Easterling et al., 1997). By way of explanation, Collatz et al. (2000) employed a simple land surface subroutine in a general circulation model of the atmosphere that included parameterizations of canopy physiological responses to various environmental changes; and by running the model with and without the vegetation subroutine, they were able to determine the degree of influence that the planet's plant life may have on near-surface air temperature in a world of rising temperature and atmospheric CO2 concentration.
In this regard, it was determined that realistic changes in the amount and physiological activity of Earth's plant life can produce changes in DTR of the order observed in the real world. In addition, the researchers explicitly state that their results "suggest that reported increases in vegetation cover in the Northern Hemisphere during the 1980s [Myneni et al., 1997] could have contributed to the lowered DTR." Hence, whereas it has long been believed that the declining diurnal temperature range near the surface of the Earth is a "fingerprint" of deleterious CO2-induced global warming, it now appears that the declining DTR may be an indication of beneficial CO2-induced "global greening," which by virtue its observation - as per Myneni et al. (1997) - is known to be helping both natural and agro-ecosystems become more productive.
In another paper, Alexandrov and Oikawa (2002) constructed a model of biospheric productivity based on empirical observations. Applied to the period 1980-90, it suggests that the total terrestrial carbon sink induced by the aerial fertilization effect of the contemporaneous increase in the air's CO2 content was approximately 1.3 Pg C yr-1, which result compares well with estimates of up to 1.1 Pg C yr-1 derived from independent empirical observations of same-period anthropogenic CO2 emissions, changes in land use, CO2 uptake by the world's oceans, and increases in the air's CO2 concentration.
Similar findings were reported nearly a decade later by Gurney and Eckles (2011). Writing as background for their report, they state that "projections of atmospheric CO2 concentrations and the resulting climate change rely to a significant degree on projections about future land and ocean uptake," citing the work of Friedlingstein et al. (2006) and Sitch et al. (2008); and, therefore, they felt it important to attempt to learn how CO2 uptake by Earth's terrestrial surfaces has varied over the past three decades. In an effort to accomplish this objective, the two authors utilized the results of atmospheric CO2 inversions - constrained by observed atmospheric CO2 concentrations (Tans et al., 1990) and simulated atmospheric transport - to estimate trends in air-to-land carbon fluxes, as per Enting (2002). This they did, as they describe it, "at spatial scales down to the continents using the results of the TransCom 3 international atmospheric CO2 inversion inter-comparison (Gurney et al., 2002, 2008)," which effort involved 13 participating modeling groups. Results of their analyses indicated that the global land carbon sink is intensifying, and that it is doing so at a rate of 0.057 PgC/year/year, resulting in 1.65 PgC of additional uptake over the period examined (1980-2008), which finding, in their words, "is consistent with related findings in recent years," citing in this regard the studies of Cao et al. (2002), Cao et al. (2005), Le Quere et al. (2009) and Piao et al. (2009).
Many researchers have also examined trends in biospheric productivity using satellite data. Nemani et al. (2003), for example, discovered a terrestrial biosphere that is growing ever more robust. Globally, the group of eight scientists determined that terrestrial net primary production (NPP) increased by 6.17%, or 3.42 PgC, over the 18 years between 1982 and 1999. What is more, they observed net positive responses over all latitude bands studied: 4.2% (47.5-22.5°S), 7.4% (22.5°S-22.5°N), 3.7% (22.5-47.5°N), and 6.6% (47.5-90.0°N).
In discussing their findings, Nemani et al. mention a number of likely contributing factors to these significant NPP increases: nitrogen deposition and forest regrowth in northern mid and high latitudes, wetter rainfall regimes in water-limited regions of Australia, Africa, and the Indian subcontinent, increased solar radiation reception over radiation-limited parts of Western Europe and the equatorial tropics, warming in many parts of the world, and the aerial fertilization effect of rising atmospheric CO2 concentrations everywhere.
With respect to the latter factor, Nemani et al. remark that "an increase in NPP of only 0.2% per 1-ppm increase in CO2 could explain all of the estimated global NPP increase of 6.17% over 18 years and is within the range of experimental evidence [italics added]." However, they report that NPP increased by more than 1% per year in Amazonia alone, noting that "this result cannot be explained solely by CO2 fertilization."
Although Nemani et al.'s conclusion may be correct, as was pointed out earlier, the aerial fertilization effect of atmospheric CO2 enrichment is most pronounced at higher temperatures, rising from next to nothing at a mean temperature of 10°C to a 0.33% NPP increase per 1-ppm increase in CO2 at a mean temperature of 36°C for a mixture of plants comprised predominantly of herbaceous species (Idso and Idso, 1994). For woody plants, this number could likely be two (Idso, 1999) or even three (Saxe et al., 1998; Idso and Kimball, 2001; Leavitt et al., 2003) times larger, yielding a 0.7% to 1% NPP increase per 1-ppm increase in CO2, which would indeed represent the lion's share of the growth stimulation observed by Nemani et al. in tropical Amazonia.
Be that as it may, the all-important take-home message of Nemani et al.'s study is that satellite-derived observations reveal the planet's terrestrial biosphere has significantly increased, as opposed to decreased, its productivity over the last two decades of the 20th century in the face of a host of both real and imagined environmental stresses.
In another study, Chen et al. (2004) utilized the monthly satellite-derived Normalized Difference Vegetation Index (NDVI) dataset of 1987-1997 that was obtained from the Advanced Very High Resolution Radiometer to calculate an 11-year history of global (75°N to 55°S) terrestrial NPP, deriving three different results based on the three different "NPP coefficient sets" of Whittaker and Likens (1975), Atjay et al. (1979) and Olson et al. (1983). Based on the beginning and end points of the graphical presentations of Chen et al.'s results, it can be calculated that global terrestrial NPP increased by approximately 6.6% between 1987 and 1997 when the Atjay et al. coefficients were used, 9.9% when the Olson et al. coefficients were used, and 13.8% when the Whittaker and Likens coefficients were used, for a mean NPP increase of about 10% over the 11-year period.
Cao et al. (2004) derived net primary production (NPP) values at 8-km and 10-day resolutions for the period 1981-2000 using variables based almost entirely on satellite observations, as described in the Global Production Efficiency Model (GLO-PEM), which consists, in their words, "of linked components that describe the processes of canopy radiation absorption, utilization, autotrophic respiration, and the regulation of these processes by environmental factors (Prince and Goward, 1995; Goetz et al., 2000)." In following this procedure, they learned that over the last two decades of the 20th century, when the heat was on, "there was an increasing trend toward enhanced terrestrial NPP," which they say was "caused mainly by increases in atmospheric carbon dioxide and precipitation."
A year later, Cao et al. (2005) used the CEVSA (Carbon Exchanges in the Vegetation-Soil-Atmosphere system) model (Cao and Woodward, 1998; Cao et al., 2002), forced by observed variations in climate and atmospheric CO2, to quantify changes in NPP, soil heterotrophic respiration (HR) and net ecosystem production (NEP) from 1981 to 1998. As an independent check on the NPP estimate of CEVSA, they also estimated 10-day NPP from 1981-2000 with the GLO-PEM model that uses data almost entirely from remote sensing, including both the normalized difference vegetation index and meteorological variables (Prince and Goward, 1995; Cao et al., 2004). This protocol revealed, in Cao et al.'s words, that "global terrestrial temperature increased by 0.21°C from the 1980s to the 1990s, and this alone increased HR more than NPP and hence reduced global annual NEP." However, they found that "combined changes in temperature and precipitation increased global NEP significantly," and that "increases in atmospheric CO2 produced further increases in NPP and NEP." They also discovered that "the CO2 fertilization effect [was] particularly strong in the tropics, compensating for the negative effect of warming on NPP." Enlarging on this point, they write that "the response of photosynthetic biochemical reactions to increases in atmospheric CO2 is greater in warmer conditions, so the CO2 fertilization effect will increase with warming in cool regions and be high in warm environments." The end result of the application of these models and measurements was their finding that global NEP increased "from 0.25 Pg C yr-1 in the 1980s to 1.36 Pg C yr-1 in the 1990s."
Commenting on their findings, Cao et al. note that "the NEP that was induced by CO2 fertilization and climatic variation accounted for 30% of the total terrestrial carbon sink implied by the atmospheric carbon budget (Schimel et al., 2001), and the fraction changed from 13% in the 1980s to 49% in the 1990s," which indicates the growing importance of the CO2 fertilization effect. Also, they say that "the increase in the terrestrial carbon sink from the 1980s to the 1990s was a continuation of the trend since the middle of the twentieth century, rather than merely a consequence of short-term climate variability," which suggests that as long as the air's CO2 content continues its upward course, so too will its stimulation of the terrestrial biosphere likely continue its upward course.
Using a newly-developed satellite-based vegetation index (Version 3 Pathfinder NDVI) in conjunction with a gridded global climate dataset (global monthly mean temperature and precipitation at 0.5° resolution from New et al., 2000), Xiao and Moody (2005) analyzed trends in global vegetative activity from 1982 to 1998. The greening trends they found exhibited substantial latitudinal and longitudinal variability, with the most intense greening of the globe located in high northern latitudes, portions of the tropics, southeastern North America and eastern China. Temperature was found to correlate strongly with greening trends in Europe, eastern Eurasia and tropical Africa. Precipitation, on the other hand, was not found to be a significant driver of increases in greenness, except for isolated and spatially fragmented regions. Some decreases in greenness were also observed, mainly in the Southern Hemisphere in southern Africa, southern South America and central Australia, which trends were associated with concomitant increases in temperature and decreases in precipitation. There were also large regions of the globe that showed no trend in greenness over the 17-year period, as well as large areas that underwent strong greening that showed no association with trends of either temperature or precipitation. These greening trends, as they concluded, must have been the result of other factors, such as "CO2 fertilization, reforestation, forest regrowth, woody plant proliferation and trends in agricultural practices."
In still another analysis, Young and Harris (2005) investigated, for the majority of Earth's land surface, a near 20-year time series (1982-1999) of NDVI data, based on measurements obtained from the Advanced Very High Resolution Radiometer (AVHRR) carried aboard U.S. National Oceanic and Atmospheric Administration satellites. In doing so, they employed two different datasets derived from the sensor: the Pathfinder AVHRR Land (PAL) data set and the Global Inventory Modeling and Mapping Studies (GIMMS) dataset. Based on their analysis of the PAL data, the two researchers determined that "globally more than 30% of land pixels increased in annual average NDVI greater than 4% and more than 16% persistently increased greater than 4%," while "during the same period less than 2% of land pixels declined in NDVI and less than 1% persistently declined." With respect to the GIMMS dataset, they report that "even more areas were found to be persistently increasing (greater than 20%) and persistently decreasing (more than 3%)." All in all, they report that "between 1982 and 1999 the general trend of vegetation change throughout the world has been one of increasing photosynthesis."
As for what has been responsible for the worldwide increase in photosynthesis - which is the ultimate food source for nearly all of the biosphere - the researchers mention global warming (perhaps it's not so bad after all), as well as "associated precipitation change and increases in atmospheric carbon dioxide," citing Myneni et al. (1997) and Ichii et al. (2002). In addition, they say that "many of the areas of decreasing NDVI are the result of human activity," primarily deforestation (Skole and Tucker, 1993; Steininger et al., 2001) and urbanization Seto et al. (2000).
One year later, Piao et al. (2006) set the stage for their study by noting that "enhanced terrestrial vegetation growth in the middle and high latitudes of the Northern Hemisphere over the past two decades has been well documented (Zhou et al., 2001; Nemani et al., 2003)," but they report that "the mechanisms for this phenomenon are still under debate." Using a leaf area index data set for the period 1981-2000, which was created from satellite-derived observations of the normalized difference vegetation index parameter for land areas above 25°N latitude, the authors therefore set out to investigate "spatial patterns of mechanisms controlling current enhanced vegetation growth in the Northern Hemisphere," focusing on "how recent changes in precipitation, temperature [and] atmospheric CO2 concentration have influenced vegetation growth." And what did they find?
Over the final two decades of the 20th century, the researchers found the mean rate of increase in growing-season leaf area index to have been 0.0041/year, and that 13% of that increase was provided by increases in precipitation, 31% was provided by increases in temperature, and that fully 49% was provided by the increase in the atmosphere's CO2 concentration.
In another study focusing on leaf area index (LAI) data derived from satellite observations, Liu et al. (2010) looked for, and computed changes in, this important plant growth parameter for six different latitude bands that included all of Earth's continents but Antarctica over the period July 1981 through December 2006. These bands were 50-90°N, 30-50°N, 10-30°N, 10°N-10°S, 10-30°S and 30-63°S. In doing so, Liu et al. determined that LAI "prominently increased" throughout Europe, Siberia, the Indian Peninsula, America and south Canada, the south region of the Sahara, the southwest corner of Australia , and the Kgalagadi Basin; while it declined in southeast Asia, southeastern China, central Africa, central and southern South America, and arctic areas in North America. Yet in spite of these latter negative results, they found that all six of the latitudinal bands they analyzed showed positive trends. Consequently, for the globe as a whole (i.e., the conglomerate of the six different latitude bands they analyzed), they determined that "LAI has increased at a rate of 0.0013 per year during July 1981-December 2006," while for the middle and high northern latitudes (north of 30°N), the linear LAI trend was 0.0032 per year.
In spite of all the positive evidences for increased global biospheric productivity listed above, some have challenged whether or not such trends are real or sustainable in a CO2-induced globally-warmed world of the future.
Introducing their work, Gloor et al. (2010) indicate that the ratio of CO2 accumulating in the atmosphere to the CO2 flux into the atmosphere due to human activities - which is known as the airborne fraction (AF) - is central to predicting changes in Earth's surface temperature due to greenhouse gas-induced warming; and they note that this ratio has remained remarkably constant over the past five decades. However, they report that Canadell et al. (2007) and Raupach et al. (2008) claim to have detected a long-term increasing trend in the airborne fraction that they interpret as being indicative of "a decreasing trend in the efficiency of the ocean and land carbon sinks."
In examining this issue, Gloor et al. report that Knorr (2009) had already challenged Canadell et al. and Raupach et al. with respect to their detection of a positive AF trend, "arguing that given the noise in the data, the trend is not detectable." Thus, they go on to challenge the second claim of Canadell et al. and Raupach et al., i.e., their contention that a positive AF trend is indicative of a decreasing planetary carbon sink efficiency by investigating "the question of what controls trends and decadal scale variations in CO2 airborne fraction using simple linear models describing the evolution of an atmospheric perturbation in CO2."
In doing so, the three researchers determined, first of all, that there is no one-to-one association between positive trends in CO2 flux to the atmosphere (due to fossil fuel emissions and changes in land use) and negative trends in Earth's carbon sink efficiency. Secondly, they found that in order to detect trends in sink efficiencies from the time course of fossil fuel-derived CO2 emissions and temporal changes in land use, "it is necessary to disentangle the spin-up time and fossil fuel growth rate variation signatures in the airborne fraction from signatures due to other causes." And when they make the pertinent calculations for fossil-fuel and land-use changes, they say they "do indeed find a positive trend in the residuals," but they argue that this trend "is not statistically significant after correcting for known events such as the temporal distribution of the extrinsic forcings and likely omissions in the emissions (particularly from land-use change)," further noting that their analysis suggests that "trends in airborne fraction are not a very good diagnostic to detect changes in carbon sink efficiency because variations in the signal are complex and the signal-to-noise ratio is small."
In describing another challenge to the recent greening of the Earth, Zhao and Running (2010) state that "previous studies have shown that climate constraints were relaxing with increasing temperature and solar radiation, allowing an upward trend in NPP [net primary production] from 1982-1999," but that over the past decade (2000-2009), satellite data "suggest a reduction in the global NPP," which finding caused some alarm among the climate-alarmist community. But, as Zhao and Running go on to show, in fact, there is likely no cause for concern.
In viewing a graphical representation of this trend (see Figure 1 below), it can be seen that apart from the starting point of the initial year (2000) of their study, there is only one other year (2004) in which the global NPP was greater than what it was at the end of the study (2009). And since global NPP was on the rise from 1982-1999, what the more recent data show would more accurately be described as a leveling off of that prior upward trend.
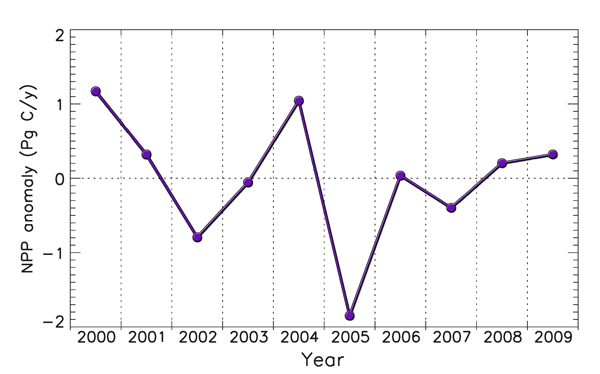
Figure 1. Interannual variations from the mean of global NPP over the past ten years. Adapted from Zhao and Running (2010).
But what about the future? Zhao and Running correctly report that the leveling off of global NPP over the past decade was induced by drought; and more frequent and more intense droughts have long been predicted by climate alarmists to accompany global warming. In this regard, therefore, it is of more than passing interest that the two researchers found that "NPP in the tropics explains 93% of variations in the global NPP," and that "tropical rainforests explain 61% of global NPP variations." These findings are especially important because of the work of Coelho and Goddard (2009), who studied El Niño-induced tropical droughts as portrayed in climate models.
In introducing their study, the latter two researchers write that "the majority of drought-related hazards and the attendant economic losses and mortality risks reside in the tropics," citing Dilley et al. (2005); and they further note that "changes in climate variability, including more frequent and damaging extreme events such as drought, is one of many anticipated impacts of climate change." More specifically (and germane to the subject at hand), they write that "El Niño brings widespread drought (i.e., precipitation deficit) to the tropics," and that "stronger or more frequent El Niño events in the future" would "exacerbate drought risk in highly vulnerable tropical areas."
As a result of these observations, the two researchers evaluated "the patterns, magnitude, and spatial extent of El Niño-induced tropical droughts during a control period in the twentieth century in climate simulations, which have realistic evolution of greenhouse gases," after which they examined "the projected changes in the characteristics of El Niño and in the strength of the identified patterns of El Niño-induced tropical drought in the twenty-first century," which then allowed them to examine patterns of mean precipitation changes in order to "assess whether those changes exacerbate or ameliorate the risk of El Niño-induced drought conditions in the twenty-first century." So what did they find?
In the first instance, Coelho and Goddard found that the models they studied "exhibit realistic patterns, magnitude, and spatial extent of El Niño-induced drought patterns in the twentieth century," and they further observed that "the teleconnections are not projected to change in the twenty-first century," although they add that "a possible slight reduction in the spatial extent of droughts is indicated over the tropics as a whole." And they report that "all model groups investigated show similar changes in mean precipitation for the end of the twenty-first century, with increased precipitation projected between 10°S and 10°N."
In conclusion, it would appear -- at least from a climate modeling perspective -- tropical drought can probably be expected to decrease throughout the remainder of the twenty-first century, which should enable the historical "greening of the earth" to continue, although there may well be decadal periods of drought-induced "leveling off" of global NPP that give rise to periodic concerns.
In the most recent satellite-based study designed to determine the net long-term trend in vegetative vigor of the entire planet, De Jong et al. (2012) employed "detection of trend changes in normalized difference vegetation index (NDVI) satellite data between 1982 and 2008," based on "time series of 648 fortnightly images [that] were analyzed using a trend breaks analysis procedure," which feat was accomplished for fourteen different classes of land cover (biomes). According to the four researchers, short-term greening and browning trends were observed over portions of the studied period for almost 15% of Earth's land surface; but they found that for the entire time period, "net greening was detected in all biomes," and that "the net global figure - considered over the full length of the time series - showed greening since the 1980s."
Finally, in one last remaining study, Ballantyne et al. (2012) used "global-scale atmospheric CO2 measurements, CO2 emission inventories and their full range of uncertainties to calculate changes in global CO2 sources and sinks during the past fifty years." In describing their results the five U.S. scientists say their mass balance analysis shows that "net global carbon uptake has increased significantly by about 0.05 billion tonnes of carbon per year and that global carbon uptake doubled, from 2.4 +/- 0.8 to 5.0 +/- 0.9 billion tonnes per year, between 1960 and 2010."
Commenting on the significance of their findings, in the concluding paragraph of their Nature article, they state that "although present predictions indicate diminished C uptake by the land and oceans in the coming century, with potentially serious consequences for the global climate, as of 2010 there is no empirical evidence that C uptake has started to diminish on the global scale." In fact, as their results clearly indicate, just the opposite appears to be the case, with global carbon uptake actually doubling over the past half-century. As such, this most recent effort refutes the recent hypothesis by Canadell et al. (2007) and Raupach et al. (2008), who claimed to have detected a decreasing trend in the efficiency of the planet's carbon sinks, and vindicates the many studies listed above revealing an ever more greening of planet Earth.
In spite of climate-alarmist claims that the temperatures of the latter part of the 20th century and on through the present were unprecedented over the past one to two millennia (which is highly debatable) and that atmospheric CO2 concentrations were the highest they had been for several hundred millennia (which is true), as well as the fact that mankind yearly harvests and/or destroys much of the planet's natural vegetation, the total yearly production of terrestrial vegetative biomass for the globe as a whole continues to rise, and at a remarkable rate. Why is this so? Perhaps it is because the twin evils of the radical environmentalist movement are not the devilish developments they are made out to be, but are actually blessings in disguise, benefiting earth's biosphere in spite of all of the political attempts to severely curtail the CO2 emissions of humanity.
References
Arft, A.M., Walker, M.D., Gurevitch, J., Alatalo, J.M., Bret-Harte, M.S., Dale, M., Diemer, M., Gugerli, F., Henry, G.H.R., Jones, M.H., Hollister, R.D., Jonsdottir, I.S., Laine, K., Levesque, E., Marion, G.M., Molau, U., Molgaard, P., Nordenhall, U., Raszhivin, V., Robinson, C.H., Starr, G., Stenstrom, A., Stenstrom, M., Totland, O., Turner, P.L., Walker, L.J., Webber, P.J., Welker, J.M. and Wookey, P.A. 1999. Responses of tundra plants to experimental warming: meta-analysis of the international tundra experiment. Ecological Monographs 69: 491-511.
Atjay, G.L., Ketner, P. and Duvigneaud, P. 1979. Terrestrial primary production and photomass. In: Bolin, B., Degens, E., Kempe, S. and Ketner, P. (Eds.), The Global Carbon Cycle, SCOPE 13, Wiley, Chichester, pp. 129-182.
Bacastow, R.B. 1976. Modulation of atmospheric carbon dioxide by the Southern Oscillation. Nature 261: 116-118.
Bacastow, R.B., Adams, J.A., Keeling, C.D., Moss, D.J., Whorf, T.P. and Wong, C.S. 1980. Atmospheric carbon dioxide, the Southern Oscillation and the weak 1975 El Niño. Science 210: 66-68.
Baker, T.R., Phillips, O.L., Malhi, Y., Almeida, S., Arroyo, L., Di Fiore, A., Erwin, T., Higuchi, N., Killeen, T.J., Laurance, S.G., Laurance, W.F., Lewis, S.L., Monteagudo, A., Neill, D.A., Núñez Vargas, P., Pitman, N.C.A., Silva, J.N.M. and Vásquez Martínez, R. 2004. Increasing biomass in Amazonian forest plots. Philosophical Transactions of the Royal Society of London Series B - Biological Sciences 359: 353-365.
Ballantyne, A.P., Alden, C.B., Miller, J.B., Tans, P.P. and White, J.W. 2012. Increase in observed net carbon dioxide uptake by land and oceans during the past 50 years. Nature 488: 70-72.
Battle, M., Bender, M.L., Tans, P.P., White, J.W.C., Ellis, J.T., Conway, T. and Francey, R.J. 2000. Global carbon sinks and their variability inferred from atmospheric O2 and 13C. Science 287: 2467-2470.
Boisvenue, C. and Running, S.W. 2006. Impacts of climate change on natural forest productivity - evidence since the middle of the 20th century. Global Change Biology 12: 862-882.
Bousquet, P., Ciais, P., Monfray, P., Balkanski, Y., Ramonet, M. and Tans, P. 1996. Influence of two atmospheric transport models on inferring sources and sinks of atmospheric CO2. Tellus Series B 48: 568-582.
Canadell, J.G., Le Quere, C., Raupach, M.R., Field, C.B., Buitenhuis, E.T., Ciais, P., Conway, T.J., Gillett, N.P., Houghton, R. and Marland, G. 2007. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proceedings of the National Academy of Sciences USA 104: 18,866-18,870.
Cao, M.K., Prince, S.D. and Shugart, H.H. 2002. Increasing terrestrial carbon uptake from the 1980s to the 1990s with changes in climate and atmospheric CO2. Global Biogeochemical Cycles 16: 10.1029/2001GB001553.
Cao, M., Prince, S.D., Small, J. and Goetz, S.J. 2004. Remotely sensed interannual variations and trends in terrestrial net primary productivity 1981-2000. Ecosystems 7: 233-242.
Cao, M., Prince, S.D., Tao, B., Small, J. and Kerang, L. 2005. Regional pattern and interannual variations in global terrestrial carbon uptake in response to changes in climate and atmospheric CO2. Tellus B 57: 210-217.
Cao, M.K. and Woodward, F.I. 1998. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature 393: 249-252.
Chen, Z.M., Babiker, I.S., Chen, Z.X., Komaki, K., Mohamed, M.A.A. and Kato, K. 2004. Estimation of interannual variation in productivity of global vegetation using NDVI data. International Journal of Remote Sensing 25: 3139-3159.
Clark, D.A. 2002. Are tropical forests an important carbon sink? Reanalysis of the long-term plot data. Ecological Applications 12: 3-7.
Clark, D.A., Piper, S.C., Keeling, C.D. and Clark, D.B. 2003. Tropical rain forest tree growth and atmospheric carbon dynamics linked to interannual temperature variation during 1984-2000. Proceedings of the National Academy of Sciences, USA 100: 10.1073/pnas.0935903100.
Coelho, C.A.S. and Goddard, L. 2009. El Niño-induced tropical droughts in climate change projections. Journal of Climate 22: 6456-6476.
Cohan, D.S., Xu, J., Greenwald, R., Bergin, M.H. and Chameides, W.L. 2002. Impact of atmospheric aerosol light scattering and absorption on terrestrial net primary productivity. Global Biogeochemical Cycles 16: 10.1029/2001GB001441.
Collatz, G.J., Bounoua, L., Los, S.O., Randall, D.A., Fung, I.Y. and Sellers, P.J. 2000. A mechanism for the influence of vegetation on the response of the diurnal temperature range to changing climate. Geophysical Research Letters 27: 3381-3384.
Condit, R. 1997. Forest turnover, density, and CO2. Trends in Ecology and Evolution 12: 249-250.
De Jong, R., Verbesselt, J., Schaepman, M.E. and De Bruin, S. 2012. Trend changes in global greening and browning: contribution of short-term trends to longer-term change. Global Change Biology 18: 642-655.
Dilley, M., Chen, R.S., Deichmann, U., Lerner-Lam, A.L. and Arnold, M. 2005. Natural Disaster Hotspots: A Global Risk Analysis. World Bank, 145 pp.
Dormann, C.F. and Woodin, S.J. 2002. Climate change in the arctic: using plant functional types in a meta-analysis of field experiments. Functional Ecology 16: 4-17.
Easterling, D.R., Horton, B., Jones, P.D., Peterson, T.C., Karl, T.R., Parker, D.E., Salinger, M.J., Razuvayev, V., Plummer, N., Jamason, P. and Folland, C.K. 1997. Maximum and minimum temperature trends for the globe. Science 277: 364-367.
Enting, I. 2002. Inverse Problems in Atmospheric Constituent Transport. Cambridge University Press, Cambridge, United Kingdom.
Esser, G., Hoffstadt, J., Mack, F. and Wittenberg, U. 1994. High Resolution Biosphere Model-Documentation of Model Version 3.00.00. Institute fur Pflanzenokologie der Justus-Liebig-Universitat, Giessen.
Feely, R.A., Wanninkhof, R., Takahashi, T. and Tans, P. 1999. Influence of El Niño on the equatorial Pacific contribution to atmospheric CO2 accumulation. Nature 398: 597-601.
Friedlingstein, P., Cox, P., Betts, R., Bopp, L., von Bloh, W., Brovkin, V., Cadule, P., Doney, S., Eby, M., Fung, I., Bala, G., John, J., Jones, C., Joos, F., Kato, T., Kawamiya, M., Knorr, W., Lindsay, K., Matthews, H.D., Raddatz, T., Rayner, P., Reick, C., Roeckner, E., Schnitzler, K.-G., Schnur, R., Strassmann, K., Weaver, A.J., Yoshikawa, C. and Zeng, N. 2006. Climate-carbon cycle feedback analysis: Results from the (CMIP)-M-4 model intercomparison. Journal of Climate 19: 3337-3353.
Gerard, J.C., Nemry, B., Francois, L.M. and Warnant, P. 1999. The interannual change of atmospheric CO2: Contribution of subtropical ecosystems? Geophysical Research Letters 26: 342-246.
Gloor, M., Sarmiento, J.L. and Gruber, N. 2010. What can be learned about carbon cycle climate feedbacks from the CO2 airborne fraction? Atmospheric Chemistry and Physics 10: 7739-7751.
Goetz, S.J., Prince, S.D., Small, J., Gleason, A.C.R. and Thawley, M.M. 2000. Interannual variability of global terrestrial primary production: reduction of a model driven with satellite observations. Journal of Geophysical Research 105: 20,007-20,091.
Grace, J., Lloyd, J., McIntyre, J., Miranda, A.C., Meir, P., Miranda, H.S., Nobre, C., Moncrieff, J., Massheder, J., Malhi, Y., Wright, I. andGash, J. 1995. Carbon dioxide uptake by an undisturbed tropical rain-forest in Southwest Amazonia, 1992-1993. Science 270: 778-780.
Graybill, D.A. and Idso, S.B. 1993. Detecting the aerial fertilization effect of atmospheric CO2 enrichment in tree-ring chronologies. Global Biogeochemical Cycles 7: 81-95.
Gu, L., Baldocchi, D., Verma, S.B., Black, T.A., Vesala, T., Falge, E.M. and Dowty, P.R. 2002. Advantages of diffuse radiation for terrestrial ecosystem productivity. Journal of Geophysical Research 107: 10.1029/2001JD001242.
Gu, L., Baldocchi, D.D., Wofsy, S.C., Munger, J.W., Urbanski, S.P. and Boden, T.A. 2003. Response of a deciduous forest to the mount Pinatubo eruption: Enhanced photosynthesis. Science 299: 2035-2038.
Gurney, K.R., Baker, D., Rayner, P., Denning, A.S. and TransCom 3 L2 modelers. 2008. Interannual variations in continental-scale net carbon exchange and sensitivity to observing networds estimated from atmospheric CO2 inversions for the period 1980 to 2005. Global Biogeochemical Cycles 22: 10.1029/2007GB003082.
Gurney, K.R., Law, R.M., Denning, A.S., Rayner, P.J., Baker, D., Bousquet, P., Bruhwiler, L., Chen, Y.-H., Ciais, P., Fan, S., Fung, I.Y., Gloor, M., Heimann, M., Higuchi, K., John, J., Maki, T., Maksyutov, S., Masarie, K., Peylin, P., Prather, M., Pak, B.C., Randerson, J., Sarmiento, J., Taguchi, S., Takahashi, T. and Yuen, C.-W. 2002. Towards robust regional estimates of CO2 sources and sinks using atmospheric transport models. Nature 415: 626-630.
Hari, P. and Arovaara, H. 1988. Detecting CO2 induced enhancement in the radial increment of trees. Evidence from the northern timberline. Scandinavian Journal of Forest Research 3: 67-74.
Hari, P., Arovaara, H., Raunemaa, T. And Hautojarvi, A. 1984. Forest growth and the effects of energy production: A method for detecting trends in the growth potential of trees. Canadian Journal of Forest Research 14: 437-440.
Houghton, R.A. 2000. Interannual variability in the global carbon cycle. Journal of Geophysical Research 105: 20,121-20,130.
Ichii, K., Kawabata, A. and Yamaguchi, Y. 2002. Global correlation analysis for NDVI and climatic variables and NDVI trends: 1982-1990. International Journal of Remote Sensing 23: 3873-3878.
Idso, C.D. and Idso, S.B. 2011. The Many Benefits of Atmospheric CO2 Enrichment. Vales Lakes Publishing, Inc., Pueblo, Colorado, USA.
Idso, K.E. and Idso, S.B. 1994. Plant responses to atmospheric CO2 enrichment in the face of environmental constraints: a review of the past 10 year's research. Agricultural and Forest Meteorology 69: 153-203.
Idso, S.B. 1986. Industrial age leading to the greening of the Earth? Nature 320: 22.
Idso, S.B. 1991a. The aerial fertilization effect of CO2 and its implications for global carbon cycling and maximum greenhouse warming. Bulletin of the American Meteorological Society 72: 962-965.
Idso, S.B. 1991b. Reply to comments of L.D. Danny Harvey, Bert Bolin, and P. Lehmann. Bulletin of the American Meteorological Society 72: 1910-1914.
Idso, S.B. 1995. CO2 and the Biosphere: The Incredible Legacy of the Industrial Revolution. Department of Soil, Water & Climate, University of Minnesota, St. Paul, Minnesota, USA.
Idso, S.B. 1999. The long-term response of trees to atmospheric CO2 enrichment. Global Change Biology 5: 493-495.
Idso, S.B. and Kimball, B.A. 2001. CO2 enrichment of sour orange trees: 13 years and counting. Environmental and Experimental Botany 46: 147-153.
Indermuhle, A., Stocker, T.F., Joos, F., Fischer, H., Smith, H.J., Wahlen, M., Deck, B., Mastroianni, D., Tschumi, J., Blunier, T., Meyer, R. and Stauffer, B. 1999. Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor Dome, Antarctica. Nature 398: 121-126.
Keeling, C.D., Whorf, T.P., Wahlen, M. and van der Pflicht, J. 1995. Interannual extremes in the rate of rise of atmospheric carbon dioxide since 1980. Nature 375: 666-670.
Knorr, W. 2000. Annual and interannual CO2 exchanges of the terrestrial biosphere: Process-based simulations and uncertainties. Global Ecology and Biogeography 9: 225-252.
Knorr, W. 2009. Is the airborne fraction of anthropogenic CO2 emissions increasing? Geophysical Research Letters 36: 10.1029/2009GL040613.
LaMarche Jr., V.C., Graybill, D.A., Fritts, H.C. and Rose, M.R. 1984. Increasing atmospheric carbon dioxide: Tree ring evidence for growth enhancement in natural vegetation. Science 223: 1019-1021.
Langenfields, R.L., Francey, R.J., Pak, B.C., Steele, L.P., Lloyd, J., Trudinger, C.M. and Allison, C.E. 2002. Interannual growth rate variations of atmospheric CO2 and its 13C, H2, CH4, and CO between 1992 and 1999 linked to biomass burning. Global Biogeochemical Cycles 16: 10.1029/2001GB001466.
Langenfelds, R.L. Francey, R.J. and Steele, L.P. 1999. Partitioning of the global fossil CO2 sink using a 19-year trend in atmospheric O2. Geophysical Research Letters 26: 1897-1900.
Law, B.E., Falge, E., Gu,. L., Baldocchi, D.D., Bakwin, P., Berbigier, P., Davis, K., Dolman, A.J., Falk, M., Fuentes, J.D., Goldstein, A., Granier, A., Grelle, A., Hollinger, D., Janssens, I.A., Jarvis, P., Jensen, N.O., Katul, G., Mahli, Y., Matteucci, G., Meyers, T., Monson, R., Munger, W., Oechel, W., Olson, R., Pilegaard, K., Paw U, K.T., Thorgeirsson, H., Valentini, R., Verma, S., Vesala, T., Wilson, K. and Wofsy, S. 2002. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agricultural and Forest Meteorology 113: 97-120.
Le Quere, C., Orr, J.C., Monfray, P., Aumont, O. and Madec, G. 2000. Interannual variability of the oceanic sink of CO2 from 1979 through 1997. Global Biogeochemical Cycles 14: 1247-1265.
Le Quere, C., Raupach, M.R., Canadell, J.G., Marland, G., Bopp, L., Ciais, P., Conway, T.J., Doney, S.C., Feely, R.A., Foster, P., Friedlingstein, P., Gurney, K., Houghton, R.A., House, J.I., Huntingford, C., Levy, P.E., Lomas, M.R., Majkut, J., Metzl, N., Ometto, J.P., Peters. G.P., Prentice, I.C., Randerson, J.T., Running, S.W., Sarmiento, J.L., Schuster, U., Sitch, S., Takahashi, T., Viovy, N., van der Werf, G.R. and Woodward, F.I. 2009. Trends in the sources and sinks of carbon dioxide. Nature Geoscience 2: 831-836.
Leavitt, S.W., Idso, S.B., Kimball, B.A., Burns, J.M., Sinha, A. and Stott, L. 2003. The effect of long-term atmospheric CO2 enrichment on the intrinsic water-use efficiency of sour orange trees. Chemosphere 50: 217-222.
Lee, K., Wanninkhof, R., Takahashi, T., Doney, S.C. and Feely, R.A. 1998. Low interannual variability in recent oceanic uptake of carbon dioxide. Nature 396: 155-159.
Lewis, S.L., Lloyd, J., Sitch, S., Mitchard, E.T.A. and Laurance, W.F. 2009. Changing ecology of tropical forests: Evidence and drivers. Annual Review of Ecology, Evolution, and Systematics 40: 529-549.
Lewis, S.L., Phillips, O.L., Baker, T.R., Lloyd, J., Malhi, Y., Almeida, S., Higuchi, N., Laurance, W.F., Neill, D.A., Silva, J.N.M., Terborgh, J., Lezama, A.T., Vásquez Martinez, R., Brown, S., Chave, J., Kuebler, C., Núñez Vargas, P. and Vinceti, B. 2004. Concerted changes in tropical forest structure and dynamics: evidence from 50 South American long-term plots. Philosophical Transactions of the Royal Society of London Series B - Biological Sciences 359: 421-436.
Lewis, S.L., Phillips, O.L., Sheil, D., Vinceti, B., Baker, T.R., Brown, S., Graham, A.W., Higuchi, N., Hilbert, D.W., Laurance, W.F., Lejoly, J., Malhi, Y., Monteagudo, A., Vargas, P.N., Sonke, B., Nur Supardi, M.N., Terborgh, J.W. and Vasquez, M.R. 2005. Tropical forest tree mortality, recruitment and turnover rates: calculation, interpretation and comparison when census intervals vary. Journal of Ecology 92: 929-944.
Lin, D., Xia, J. and Wan, S. 2010. Climate warming and biomass accumulation of terrestrial plants: a meta-analysis. New Phytologist 188: 187-198.
Liu, S., Liu, R. and Liu, Y. 2010. Spatial and temporal variation of global LAI during 1981-20006. Journal of Geographical Sciences 20: 323-332.
Malhi Y. and Grace, J. 2000. Tropical forests and atmospheric carbon dioxide. Trends in Ecology and Evolution 15: 332-337.
Malhi, Y., Nobre, A.D., Grace, J., Kruijt, B., Pereira, M.G.P., Culf, A. And Scott, S. 1998. Carbon dioxide transfer over a Central Amazonian rain forest. Journal of Geophysical Research 103: 31,593-31,612.
Myneni, R.C., Keeling, C.D., Tucker, C.J., Asrar, G. and Nemani, R.R. 1997. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 386: 698-702.
Nemani, R.R., Keeling, C.D., Hashimoto, H., Jolly, W.M., Piper, S.C., Tucker, C.J., Myneni, R.B. and Running. S.W. 2003. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300: 1560-1563.
Olson, J.S., Watts, J. and Allison, L. 1983. Carbon in Live Vegetation of Major World Ecosystems, Environmental Sciences Division, Oak Ridge National Laboratory, Oak Ridge, TN, USA.
Parker, M.L. 1987. Recent abnormal increase in tree-ring widths: A possible effect of elevated atmospheric carbon dioxide. In: Jacoby Jr., G.C. and Hornbeck, J.W. (Eds.), Proceedings of the International Symposium on Ecological Aspects of Tree-Ring Analysis. U.S. Department of Energy, Washington, DC, pp. 511-521.
Phillips, O.L., Baker, T.R., Arroyo, L., Higuchi, N., Killeen, T.J., Laurance, W.F., Lewis, S.L., Lloyd, J., Malhi, Y., Monteagudo, A., Neill, D.A., Núñez Vargas, P., Silva, J.N.M., Terborgh, J., Vásquez Martínez, R., Alexiades, M., Almeida, S., Brown, S., Chave, J., Comiskey, J.A., Czimczik, C.I., Di Fiore, A., Erwin, T., Kuebler, C., Laurance, S.G., Nascimento, H.E.M., Olivier, J., Palacios, W., Patiño, S., Pitman, N.C.A., Quesada, C.A., Saldias, M., Torres Lezama, A., B. and Vinceti, B. 2004. Pattern and process in Amazon tree turnover: 1976-2001. Philosophical Transactions of the Royal Society of London Series B - Biological Sciences 359: 381-407.
Phillips, O.L. and Gentry, A.H. 1994. Increasing turnover through time in tropical forests. Science 263: 954-958.
Phillips, O.L., Malhi, Y., Higuchi, N., Laurance, W.F., Nunez, P.V., Vasquez, R.M., Laurance, S.G., Ferreira, L.V., Stern, M., Brown, S. and Grace, J. 1998. Changes in the carbon balance of tropical forests: Evidence from long-term plots. Science 282: 439-442.
Phillips, O.L., Malhi, Y., Vinceti, B., Baker, T., Lewis, S.L., Higuchi, N., Laurance, W.F., Vargas, P.N., Martinez, R.V., Laurance, S., Ferreira, L.V., Stern, M., Brown, S. and Grace, J. 2002. Changes in growth of tropical forests: Evaluating potential biases. Ecological Applications 12: 576-587.
Piao, S., Ciais, P., Friedlingstein, P., de Noblet-Ducoudre, N., Cadule, P., Viovy, N. and Wang, T. 2009. Spatiotemporal patterns of terrestrial carbon cycle during the 20th century. Global Biogeochemical Cycles 23: 10.1029/2008GB003339.
Piao, S., Friedlingstein, P., Ciais, P., Zhou, L. and Chen, A. 2006. Effect of climate and CO2 changes on the greening of the Northern Hemisphere over the past two decades. Geophysical Research Letters 33: 10.1029/2006GL028205.
Pimm, S.L. and Sugden, A.M. 1994. Tropical diversity and global change. Science 263: 933-934.
Prentice, I.C., Farquhar, G.D., Fasham, M.J.R., Goulden, M.L., Heimann, M., Jaramillo, V.J., Kheshgi, H.S., Le Quere, C., Scholes, R.J., Wallace, D.W.R., Archer, D., Ashmore, M.R., Aumont, O., Baker, D., Battle, M., Bender, M., Bopp, L.P., Bousquet, P., Caldeira, K., Ciais, P., Cox, P.M., Cramer, W., Dentener, F., Enting, I.G., Field, C.B., Friedlingstein, P., Holland, E.A., Houghton, R.A., House, J.I., Ishida, A., Jain, A.K., Janssens, I.A., Joos, F., Kaminski, T., Keeling, C.D., Keeling, R.F., Kicklighter, D.W., Hohfeld, K.E., Knorr, W., Law, R., Lenton, T., Lindsay, K., Maier-Reimer, E., Manning, A.C., Matear, R.J., McGuire, A.D., Melillo, J.M., Meyer, R., Mund, M., Orr, J.C., Piper, S., Plattner, K., Rayner, P.J., Sitch, S., Slater, R., Taguchi, S., Tans, P.P., Tian, H.Q., Weirig, M.F., Whorf, T. and Yool, A. 2001. The carbon cycle and atmospheric carbon dioxide. Chapter 3 of the Third Assessment Report of the Intergovernmental Panel on Climate Change. Climate Change 2001: The Scientific Basis. Cambridge University Press, Cambridge, UK, pp. 183-238.
Prince, S.D. and Goward, S.N. 1995. Global primary production: a remote sensing approach. Journal of Biogeography 22: 815-835.
Raupach, M.R., Canadell, J.G. and Le Quere, C. 2008. Anthropogenic and biophysical contributions to increasing atmospheric CO2 growth rate and airborne fraction. Biogeosciences 5: 1601-1613.
Rayner, P.J. and Law, R.M. 1999. The interannual variability of the global carbon cycle. Tellus Series B 51: 210-212.
Reichenau, T.G. and Esser, G. 2003. Is interannual fluctuation of atmospheric CO2 dominated by combined effects of ENSO and volcanic aerosols? Global Biogeochemical Cycles 17: 10.1029/2002GB002025.
Roderick, M.L., Farquhar, G.D., Berry, S.L. and Noble, I.R. 2001. On the direct effect of clouds and atmospheric particles on the productivity and structure of vegetation. Oecologia 129: 21-30.
Saxe, H., Ellsworth, D.S. and Heath, J. 1998. Tree and forest functioning in an enriched CO2 atmosphere. New Phytologist 139: 395-436.
Schimel, D.S., House, J.I., Hibbard, J.I., Bousquet, P., Ciais, P., Peylin, P., Braswell, B.H., Apps, M.J., Baker, D., Bondeau, A., Canadell, J., Churkina, G., Cramer, W., Denning, A.S., Field, C.B., Friedlingstein, P., Goodale, C., Heimann, M., Houghton, R.A., Melillo, J.M., Moore III, B., Murdiyarso, D., Noble, I., Pacala, S.W., Prentice, I.C., Raupach, M.R., Rayner, P.J., Scholes, R.J., Steffen, W.L. and Wirth, C. 2001. Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature 414: 169-172.
Seto, K.C., Kaufman, R.K. and Woodcock, C.E. 2000. Landsat reveals China's farmland reserves, but they're vanishing fast. Nature 406: 121.
Sheil, D. 1995. Evaluating turnover in tropical forests. Science 268: 894.
Sheil, D. and May, R.M. 1996. Mortality and recruitment rate evaluations in heterogeneous tropical forests. Journal of Ecology 84: 91-100.
Sitch, S., Huntingford, C., Gedney, N., Levy, P.E., Lomas, M., Piao, S.L., Betts, R., Ciais, P., Cox, P., Friedlingstein, P., Jones, C.D., Prentice, I.C. and Woodward, F.I. 2008. Evaluation of the terrestrial carbon cycle, future plant geography and climate-carbon cycle feedbacks using five Dynamic Global Vegetation Models (DGVMs). Global Change Biology 14: 2015-2039.
Skole, D. and Tucker, C.J. 1993. Tropical deforestation and habitat fragmentation in the Amazon: satellite data from 1978 to 1988. Science 260: 1905-1909.
Steininger, M.K., Tucker, C.J., Ersts, P., Killen, T.J., Villegas, Z. and Hecht, S.B. 2001. Clearance and fragmentation of tropical deciduous forest in the Tierras Bajas, Santa Cruz, Bolivia. Conservation Biology 15: 856-866.
Tans, P.P., Fung, I.Y. and Takahashi, T. 1990. Observational constraints on the global atmospheric CO2 budget. Science 247: 1431-1438.
Walker, M.D., Wahren, C.H., Hollister, R.D., Henry, G.H.R., Ahlquist, L.E., Alatalo, J.M., Bret-Harte, M.S., Calef, M.P., Callaghan, T.V., Carroll, A.B., Epstein, H.E., Jonsdottir, I.S., Klein, J.A., Magnusson, B., Molaug, U., Oberbauer, S.F., Rewan, S.P., Robinson, C.H., Shaver, G.R., suding, K.N., Thompson, C.C., Tolvanen, A., Totland, O., Turner, P.L., Tweedie, C.E., Webber, P.J. and Wookey, P.A. 2006. Plant community responses to experimental warming across the tundra biome. Proceedings of the National Academy of Sciences USA 103: 1342-1346.
Weaver, P.L. and Murphy, P.G. 1990. Forest structure and productivity in Puerto Rico's Luquillo Mountains. Biotropica 22: 69-82.
West, D.C. 1988. Detection of forest response to increased atmospheric carbon dioxide. In: Koomanoff, F.A. (Ed.), Carbon Dioxide and Climate: Summaries of Research in FY 1988. U.S. Department of Energy, Washington, D.C., p. 57.
Whittaker, R.H. and Likens, G.E. 1975. The biosphere and man. In: Leith, H. and Whittaker, R.H. (Eds.), Primary Productivity and the Biosphere, Ecological Studies 14, Springer-Verlag, Berlin, Germany, pp. 305-328.
Wittwer, S.H. 1995. Food, Climate, and Carbon Dioxide: The Global Environment and World Food Production. Lewis Publishers, Boca Raton, FL.
Xiao, J. and Moody, A. 2005. Geographical distribution of global greening trends and their climatic correlates: 1982-1998. International Journal of Remote Sensing 26: 2371-2390.
Young, S.S. and Harris, R. 2005. Changing patterns of global-scale vegetation photosynthesis, 1982-1999. International Journal of Remote Sensing 26: 4537-4563.
Zhou, L., Tucker, C.J., Kaufmann, R.K., Slayback, D., Shabanov, N.V. and Myneni, R.B. 2001. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981-1999. Journal of Geophysical Research 106: 20,069-20,083.
Zhao, M. and Running, S.W. 2010. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 329: 940-943.
Last updated 3 September 2014