




Among the many alarmist fears of CO2-induced global warming is the concern that the productivity of the biosphere will decline if global temperatures rise to the extent predicted by computer models. Because of this concern, several researchers have investigated the relationship between temperature, atmospheric CO2, and biospheric productivity across a range of spatial and temporal scales. The present review examines what those researchers have learned, focusing on studies that have addressed this issue for the globe as a whole over both the distant and historic past.
Several years ago, based on a series of empirically-derived mathematical relationships, Luz et al. (1999) developed a model of the isotopic composition of oxygen trapped in air bubbles in a Greenland ice core that they used to estimate global biospheric productivity over the past 82,000 years. This work suggested that biospheric productivity at 18,000, 26,000, 37,000, 56,000 and 82,000 years before present was, respectively, 89%, 91%, 87%, 97% and 91% of what it was approximately 150 years prior to the time of their study, when the atmospheric CO2 concentration was approximately 280 ppm.
These results imply that biospheric productivity a century and a half ago was at its highest point of the past 82,000 years, as was the atmosphere's CO2 concentration. Therefore, assuming the air's CO2 content is the primary determinant of biospheric productivity, and based on the mean result for the five periods Luz et al. investigated, it can be calculated that the 73-ppm increase (from 207 to 280 ppm) in the air's CO2 concentration between the earlier period and 1850 was responsible for a 9.9% increase (from the earlier/later productivity ratio change of 0.91 to 1.00) in planet-wide biospheric productivity. And assuming a linear relationship between these two parameters, it can be calculated that a 300-ppm increase in atmospheric CO2 concentration would produce an approximate 40% increase in biospheric productivity, which is essentially the mean value of the CO2-growth response that has been derived from the thousands of studies that have evaluated this phenomenon for hundreds of Earth's plants (see the CO2 Science Plant Growth Database).
In a contemporaneous study, but involving a much shorter timescale, Indermuhle et al. (1999) examined the age and composition of air bubbles trapped in glacial ice cores retrieved from Taylor Dome, Antarctica, which enabled them to reconstruct a history of carbon exchanges among the atmosphere, oceans and land biota over the past 11,000 years. This effort indicated that the atmosphere's CO2 concentration at approximately 10,500 years before present (yr BP) was 268 ppm, that it fell to a value of 260 ppm at about 8,200 yr BP, and that it increased monotonically and "almost linearly" from that point in time to a value of 285 ppm 7,000 years later (about 1,200 yr BP). Model calculations based on carbon isotope data further revealed that the drawdown of atmospheric CO2 from 10,500 to 8,200 yr BP was consistent with terrestrial vegetative regrowth and soil build-up on areas previously covered by ice sheets, "as well as a climatic development towards the mid-Holocene optimum." Thereafter, however, the scientists' work revealed a gradual loss of terrestrial carbon over the next 7,000 years, which they say was likely "due to a change from the warmer and wetter mid-Holocene climate to colder and drier conditions."
These observations confirm that the warm and moist mid-Holocene has not been called the Holocene Climatic Optimum for nothing. Indeed, Indermuhle et al.'s work convincingly shows that it was the premier period of terrestrial vegetative prowess of the present interglacial, and that Earth's biosphere has been going steadily downhill ever since (or at least to the start of the Industrial Revolution). Even the 25 ppm increase in the air's CO2 content from 8,200 to 1,200 yr BP was not enough to stem the downward biological spiral induced by the planet's slowly deteriorating climate. Now, however, with an anthropogenically-induced rate of increase in atmospheric CO2 concentration that is fully two orders of magnitude greater, this situation will likely reverse itself, as it has indeed been doing ever since the demise of the Little Ice Age.
In another study of the same general time period, Ward et al. (2005) compared "the physiological (stomatal-regulation) and modeled-growth responses of glacial and modern Juniperus trees ... by using stable carbon isotope methodologies," working with what they describe as "a very rare series of preserved Juniperus (C3) wood specimens that span a large portion of the last glacial period." Values of both carbon isotope discrimination and the ratio of internal leaf to atmospheric CO2 concentration (ci/ca) at the time of the last glacial maximum were determined to be the same as those of today, implying that "leaves of full-glacial trees had extremely low calculated ci values (averaging 113 ppm) that were 25% lower than in leaves of postglacial trees (ci of 150 ppm between 7.665 and 12.450 kyr B.P.), and 40% lower than in leaves of modern trees (average ci of 187 ppm)."
Noting that "previous studies with modern C3 plants indicate that ci values are highly scaleable to photosynthetic rate and growth, with particularly strong correlations occurring at low CO2 conditions (Polley et al., 1993; Sage and Coleman, 2001)," Ward et al. estimated that forest productivity was reduced by about 55% between modern and full-glacial ci values. Viewed from the perspective of forward-evolving history, this finding suggests that most of Earth's trees have already seen their productivity more than doubled by the aerial fertilization effect of the increase in the air's CO2 concentration experienced over the past 12.5 thousand years.
Pointing out that the low Last Glacial Maximum ci values they found "were not unique to southern California, because glacial leaves of Pinus flexilis from the Great Basin exhibited ci values of 110 ppm (van de Water et al., 1994), supporting the notion that trees in nearby regions were also carbon-starved during the last glacial period," Ward et al. ultimately concluded that "the productivity of ancient trees, along with other C3 species, was greatly diminished during the last glacial period." The words carbon-starved are their choice for describing this situation, and they appear in several places throughout their paper. And given such findings, it is clear that the concomitant increases in atmospheric CO2 concentration and air temperature experienced from the Last Glacial Maximum to the present have truly been a godsend for the productivity of the biosphere.
Over the more recent past, other studies have produced additional evidence for a worldwide increase in vegetative productivity since the inception of the Industrial Revolution. Joos and Bruno (1998), for example, used ice core data and direct observations of atmospheric CO2 and 13C to reconstruct the histories of terrestrial and oceanic uptake of anthropogenic carbon over the past two centuries. In doing so, they discovered the biosphere typically acted as a source of CO2 during the 19th century and the first decades of the 20th century, but that it subsequently "turned into a sink." In another paper, Lloyd (1999) calculated that from 1730 to the early 1980s the increase in temperate deciduous forest net primary productivity (NPP) due solely to the historical increase in the atmosphere's CO2 concentration was approximately 7%, and that the increase in NPP due to a modest proportional increase in nitrogen deposition over the same time period would have been about 25%. However, when CO2 and nitrogen increased together in the model employed by Lloyd, the NPP stimulation was 40%, which is more than the sum of the individual contributions of the extra CO2 and nitrogen.
In a subsequent study, Tans (2009) employed measurements of atmospheric and oceanic carbon contents, along with reasonably constrained estimates of global anthropogenic CO2 emissions, to calculate the residual fluxes of carbon (in the form of CO2) from the terrestrial biosphere to the atmosphere (+) or from the atmosphere to the terrestrial biosphere (-), obtaining the results depicted in the figure below.
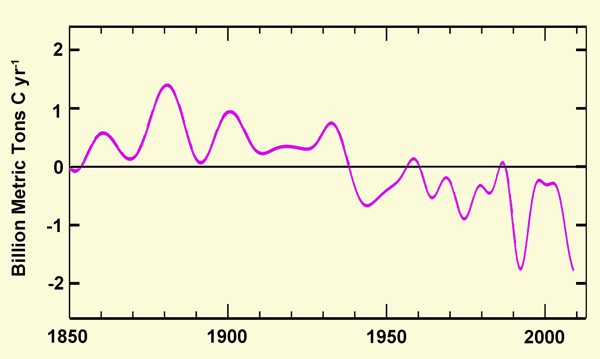
Five-year smoothed rates of carbon transfer from land to air (+) or from air to land (-) vs. time. Adapted from Tans (2009).
As can be seen from this figure, Earth's land surfaces were a net source of CO2-carbon to the atmosphere until about 1940, primarily due to the felling of forests and the plowing of grasslands to make way for expanded agricultural activities. From 1940 onward, however, the terrestrial biosphere has become, in the mean, an increasingly greater sink for CO2-carbon; and it has done so even in the face of massive global deforestation, for which it has more than compensated.
Such findings, which do "not depend on models" but "only on the observed atmospheric increase and estimates of fossil fuel emissions," led Tans to conclude that "suggestions that the carbon cycle is becoming less effective in removing CO2 from the atmosphere (e.g., LeQuere et al., 2007; Canadell et al., 2007) can perhaps be true locally, but they do not apply globally, not over the 50-year atmospheric record, and not in recent years." In fact, Tans goes on to say that "to the contrary" and "despite global fossil fuel emissions increasing from 6.57 GtC in 1999 to 8.23 in 2006, the five-year smoothed global atmospheric growth rate has not increased during that time, which requires more effective uptake [of CO2] either by the ocean or by the terrestrial biosphere, or both, to satisfy atmospheric observations." And the results portrayed in the figure above that was adapted from Tans' paper clearly indicate that this "more effective uptake" of CO2-carbon has occurred primarily over land.
Lastly, in a study that did depend on models, Qian et al. (2010) analyzed the relationship between CO2, temperature, and biospheric productivity using ten different models that took part in the Coupled Carbon Cycle Climate Model Intercomparison Project (C4MIP) of the International Geosphere-Biosphere Program and the World Climate Research Program, all of which models, in their words, "used the same anthropogenic fossil fuel emissions from Marland et al. (2005) from the beginning of the industrial period until 2000 and the IPCC SRES A2 scenario for the 2000-2100 period." In doing so, they learned that the ten C4MIP models predicted a mean warming of 5.6°C from 1901 to 2100 in the Earth's Northern High Latitudes (NHLs, land poleward of 60°N); and the three researchers state that "the NHL will be a carbon sink of 0.3 +/- 0.3 PgCyr-1 by 2100." They also state that "the cumulative land organic carbon storage is modeled to increase by 38 +/- 20 PgC over 1901 levels, of which 17 +/- 8 PgC comes from vegetation [a 43% increase] and 21 +/- 16 PgC from the soil [an 8% increase]," noting that "both CO2 fertilization and warming enhance vegetation growth in the NHL." Such findings suggest that the warming experienced during the 20th century, coupled with the warming projected by the models to occur through the end of the 21st century, has not been - nor will it likely be - a detriment to plant growth and productivity in the NHL.
In conclusion, in spite of claims that rising atmospheric CO2 concentrations and unprecedented global warming since the inception of the Industrial Revolution are destroying (or will destroy) the productivity of the biosphere, the terrestrial vegetative biomass of the globe as a whole continues to rise; and it appears to be doing so at a remarkable rate. As for why is this so, it may well be that the twin evils of the radical environmental movement (rising temperatures and atmospheric CO2 concentrations) are not the evils they are made out to be, but actually blessings in disguise ... blessings that are fueling the biosphere!
References
Canadell, J.G., LeQuere, C., Raupach, M.R., Field, C.B., Buitenhuis, E.T,., Ciais, P., Conway, T.J., Gillett, N.P., Houghton, R.A. and Marland, G. 2007. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proceedings of the National Academy of Sciences of the United States of America 104: 18,866-18.870.
Indermuhle, A., Stocker, T.F., Joos, F., Fischer, H., Smith, H.J., Wahlen, M., Deck, B., Mastroianni, D., Tschumi, J., Blunier, T., Meyer, R. and Stauffer, B. 1999. Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor Dome, Antarctica. Nature 398: 121-126.
Joos, F. and Bruno, M. 1998. Long-term variability of the terrestrial and oceanic carbon sinks and the budgets of the carbon isotopes 13C and 14C. Global Biogeochemical Cycles 12: 277-295.
LeQuere, C., Rodenbeck, C., Buitenhuis, E.T., Conway, T.J., Langenfelds, R., Gomez, A., Labuschagne, C., Ramonet, M., Nakazawa, T., Metzl, N., Gillett, N. and Heimann, M. 2007. Saturation of the Southern Ocean CO2 sink due to recent climate change. Science 316: 1735-1738.
Lloyd, J. 1999. The CO2 dependence of photosynthesis, plant growth responses to elevated CO2 concentrations and their interaction with soil nutrient status, II. Temperate and boreal forest productivity and the combined effects of increasing CO2 concentrations and increased nitrogen deposition at a global scale. Functional Ecology 13: 439-459.
Luz, B., Barkan, E., Bender, M.L., Thiemens, M.H. and Boering, K.A. 1999. Triple-isotope composition of atmospheric oxygen as a tracer of biospheric productivity. Nature 400: 547-550.
Marland, G., Boden, T.A. and Andres, R.J. 2005. Global, regional, and national CO2 emissions. In: Trends: A Compendium of Data on Global Change. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy, Oak Ridge, Tennessee, USA. Available at http://cdiac.ornl.gov/trends/emis/overview.html.
Polley, H.W., Johnson, H.B., Marino, B.D. and Mayeux, H.S. 1993. Increases in C3 plant water-use efficiency and biomass over glacial to present CO2 concentrations. Nature 361: 61-64.
Qian, H., Joseph, R. and Zeng, N. 2010. Enhanced terrestrial carbon uptake in the Northern High Latitudes in the 21st century from the Coupled Carbon Cycle Climate Model Intercomparison Project model projections. Global Change Biology 16: 641-656.
Sage, R.F. and Coleman, J.R. 2001. Effects of low atmospheric CO2 on plants: more than a thing of the past. Trends in Plant Science 6: 323-326.
Tans, P. 2009. An accounting of the observed increase in oceanic and atmospheric CO2 and an outlook for the future. Oceanography 22: 26-35.
Van de Water, P.K., Leavitt, S.W. and Betancourt, J.L. 1994. Trends in stomatal density and 13C/12C ratios of Pinus flexilis needles during the last glacial-interglacial cycle. Science 264: 239-243.
Ward, J.K., Harris, J.M., Cerling, T.E., Wiedenhoeft, A., Lott, M.J., Dearing, M.-D., Coltrain, J.B. and Ehleringer, J.R. 2005. Carbon starvation in glacial trees recovered from the La Brea tar pits, southern California. Proceedings of the National Academy of Sciences, USA 102: 690-694.
Last updated 4 April 2012