




The State of Earth's Terrestrial Biosphere:
How is it Responding to Rising Atmospheric CO2 and Warmer Temperatures?
Continental-Scale Analyses of Terrestrial Productivity: North America - High Latitude Regions
For many years, the story out of the Arctic was that rising temperatures would change the land from a carbon sink to a carbon source, further exacerbating the cause of ecosystem change, i.e., regional warming, by adding to the atmosphere's burden of greenhouse gases and hastening that portion of the biosphere's inevitable degradation (Oechel et al., 1993, 1995). In the early to mid-1970s, for example, when the first carbon balance studies of Alaskan Arctic ecosystems were conducted, both wet-sedge communities and moist-tussock tundra were observed to be net sinks of carbon. By the mid-1980s and early 1990s, however, following significant increases in air temperature and surface water deficit, both ecosystems had become net sources of carbon. Then, between 1992 and 1996, in response to further warming and drying that resulted, in the words of Oechel et al. (2000a), in "the highest average summer temperature and surface water deficit observed for the entire 39-year period," both ecosystems' net summer releases of CO2 to the atmosphere declined, and they eventually became CO2 sinks.
How did it happen? In the words of the scientists who conducted the work, their observations simply indicated "a previously undemonstrated capacity for ecosystems to metabolically adjust to long-term (decadal or longer) changes in climate."
But how did that happen? Was there help along the way from the concomitant rise in the air's CO2 content and its aerial fertilization and anti-transpirant effects? Although these well-documented consequences of atmospheric CO2 enrichment are known to enable plants to better respond to the environmental challenges of both warming and drying, these effects were downplayed. Instead, the researchers noted some other possibilities that are also plausible.
First, there is the likelihood that during the initial stages of warming and soil drying, younger and more labile carbon would be rapidly decomposed, shifting the net summer carbon balance of the ecosystems from one of carbon sequestration to one of carbon evolution. After this initial perturbation, however, Oechel et al. suggest that "enhanced rates of net nitrogen-mineralization should eventually stimulate rates of gross primary production and atmospheric CO2 sequestration."
Another possibility is a gradual shift in plant species towards more productive types that would further reduce the large initial carbon losses over time. Writing on this subject, the researchers say "there is evidence that the relative abundance of deciduous shrubs has increased in response to climate change over the past 1-2 decades in Alaskan moist-tussock tundra ecosystems," which is also something that is expected to occur as a consequence of the ongoing rise in the air's CO2 content. Capers and Stone (2011), for example, have reported that in response to rising air temperatures, "trees have established where they did not previously occur, both in alpine areas (Wardle and Coleman, 1992; Peterson, 1994; Kullman, 2001, 2002), and in arctic tundra (Lescop-Sinclair and Payette, 1995; Danby and Hik, 2007)," while "increasing shrub abundance also has been reported in alpine (Klanderud and Birks, 2003) and arctic locations (Sturm et al., 2001; Tape et al., 2006; Wilson and Nilsson, 2009)." Be that as it may, and whatever the cause (or causes), it is clear that the productivity of vegetation in the northern reaches of the Northern Hemisphere has been increasing with time, as further illustrated in the following studies.
Noting that "canopy-forming shrubs are reported to be increasing at sites around the circumpolar Arctic," Myers-Smith et al. (2011) (1) examined historic photographs, (2) repeated vegetation surveys, and (3) conducted monitoring of long-term plots on Canada's Herschel Island (69.57°N, 138.91°W), in order to see what had occurred over the past century on this 100-km2 parcel of land just off the western Arctic coast of Canada's Yukon Territory. And based on all three of their lines of research, they say they "found evidence of increases of each of the dominant canopy-forming willow species (Salix richardsonii, Salix glauca and Salix pulchra), during the twentieth century," along with evidence that "the majority of willow patches for each of these species became established between 1910 and 1960," but "with stem ages and maximum growth rates indicating that some patches could have established as late as the 1980s."
In discussing their findings, the seven scientists report that they represent but one example of the 20th-century greening of the Circumpolar Arctic, noting that "recent evidence indicates an expansion of canopy-forming shrubs at sites on the North Slope of Alaska (Sturm et al., 2001; Tape et al., 2006), on the coast of the Northwest Territories (Lantz et al., 2009), in Northern Quebec (B. Tremblay et al., personal communication), and in northern Russia (Forbes et al., 2010a)." In addition, they indicate that "in Arctic Alaska, canopy cover of alder shrubs has increased by 14-20% on average within the last 40 years, with increases of up to 80% in some areas (Tape et al., 2006)." And they say that "studies of population structures of shrub and tree species indicate advancing of shrubs up slopes in alpine tundra ecosystems in subarctic Sweden (Hallinger et al., 2010)," as well as in sites in Norway (Hofgaard et al., 2009), while further noting that "local indigenous Nenets people in the western Russian Arctic report increasing willow shrubs (Forbes et al., 2010b)" and that "similar observations of vegetation change by Inuit have been reported in Arctic Canada (Thorpe et al., 2002)."
It is also of interest to note that the team of Canadian researchers states that "pollen records indicate that willows were widespread in Arctic ecosystems during warmer periods after the last glacial maximum (Brubaker et al., 1983; Bigelow et al. (2003)." And these several observations would seem to suggest that the entire Circumpolar Arctic is in the process of returning to what could be called the good old days, when that part of the planet was a whole lot greener - and a whole lot livelier - than it has been for a long, long time.
Also documenting increased plant growth in the high latitudes of North America was Goetz et al. (2005), who transformed satellite-derived NDVI data obtained across boreal North America (Canada and Alaska) for the period 1982-2003 into photosynthetically-active radiation absorbed by green vegetation and treated the result as a proxy for relative June-August gross photosynthesis (Pg), stratifying the results by vegetation type and comparing them with spatially-matched concomitant trends in surface air temperature data. Over the course of the study, this work revealed that area-wide tundra experienced a significant increase in Pg in response to a similar increase in air temperature; and Goetz et al. say that "this observation is supported by a wide and increasing range of local field measurements characterizing elevated net CO2 uptake (Oechel et al., 2000b), greater depths of seasonal thaw (Goulden et al., 1998), changes in the composition and density of herbaceous vegetation (Chapin et al., 2000; Epstein et al., 2004), and increased woody encroachment in the tundra areas of North America (Sturm et al., 2001)." In the case of interior forest, on the other hand, there was no significant increase in air temperature and essentially no change in Pg, with the last data point of the series being essentially indistinguishable from the first. This latter seemingly aberrant observation is in harmony with the fact that at low temperatures the growth-promoting effects of increasing atmospheric CO2 levels are often very small or even non-existent (Idso and Idso, 1994), which is what appears to have been the case with North American boreal forests over the same time period. As a result, Canada's and Alaska's tundra ecosystems exhibited increasing productivity over the past couple of decades, while their boreal forests did not.
Writing as background for their study, Ropars and Boudreau (2012) state that "Myneni et al. (1997, 1998) were the first to report evidence of the pan-Arctic increase in vegetation cover," noting that "by analyzing worldwide NDVI [Normalized Difference Vegetation Index] trends between 1981 and 1991, they showed that the greatest increase in photosynthetic activity occurred in regions above 50°N," and that "since then, this phenomenon has been observed for different regions," including Alaska (Silapaswan et al., 2001; Jia et al., 2003; Verbyla, 2008), Russia (Forbes et al., 2010a), Western Canada (Olthof and Pouliot, 2010), over a longer time span (Jia et al., 2003; Goetz et al., 2005; Verbyla, 2008; Forbes et al., 2010a; Olthof and Pouliot, 2010) and at a better resolution (Jia et al., 2003; Olthof and Pouliot, 2010). Against this backdrop and focusing on an area near the Boniface River research station 35 km east of Hudson Bay and 10 km south of the treeline in subarctic Quebec (Canada), Ropars and Boudreau (2012) evaluated changes in shrub cover over a full half-century, via their comparison of two aerial photographs taken in July 1957 and a satellite image of the same area obtained in July 2008.
The efforts of the two researchers revealed that "both hilltops and terraces recorded an increase in shrub cover" (Figure 20), and they say that "the increase was significantly greater on terraces than on hilltops (21.6% versus 11.6%)," while further noting that "this finding corroborates other studies using a similar method conducted in different regions of the Arctic," including Alaska (Sturm et al., 2001; Tape et al., 2006), northern Quebec (Tremblay, 2010), Russia (Forbes et al., 2010) and studies which revealed a major increase of the NDVI over the last few decades (Jia et al., 2003; Verbyla, 2008). Ropars and Boudreau conclude their report by stating that "according to ground truthing, the shrub cover densification is associated mainly with an increase of Betula glandulosa Michx" - more commonly known as dwarf birch - and by noting that "the numerous seedlings observed during the ground truthing suggest that shrub densification should continue in the future."
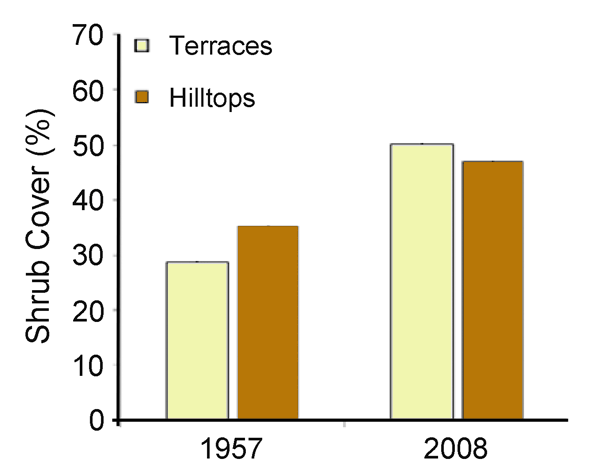
Figure 20. Percent shrub cover on terraces and hilltops in subarctic Quebec, Canada in 1957 and 2008. Adapted from Ropars and Boudreau (2012).
In the introduction to their report on the response of High Arctic tundra vegetation to the warming experienced in that part of the world over the past quarter-century, Hudson and Henry (2009) note that the Arctic has warmed by about 1.6°C over the past four decades, citing the analysis of McBean et al. (2005); and they state that this temperature increase has "led the Arctic Climate Impact Assessment (ACIA) and Intergovernmental Panel on Climate Change (IPCC) to predict that tundra ecosystems will be particularly threatened by climate change [i.e., warming] over the next century." But are such predictions correct?
In an effort to provide some real-world perspective on the issue, Hudson and Henry measured biomass and composition changes in a heath community dominated by several vascular plants and bryophytes at an 8-km2 coastal lowland adjacent to Alexandra Fiord on the east-central coast of Ellesmere Island, Nunavut, Canada. This they did over a period of 13 years (1995-2007), using a point-intercept method in permanent plots, and over a period of 27 years (1981-2008) using a biomass harvest comparison. And what did they find?
"Results from both methods," in the words of the Canadian scientists, "indicate that the community became more productive over time," that "bryophyte and evergreen shrub abundances increased," while "deciduous shrub, forb, graminoid, and lichen cover did not change," so that "species diversity also remained unchanged [italics added]," all of which changes - and non-changes -- stand in stark contrast to the "particularly threatening" view of the ACIA and IPCC.
In further support of their "contrary" findings, Hudson and Henry additionally report that "satellite-based remote sensing models, such as green trends derived from the normalized difference vegetation index (NDVI; e.g., Myneni et al., 1997; Zhou et al., 2001; Stow et al., 2004; Verbyla, 2008), and global vegetation and ecosystem process simulations of the terrestrial carbon cycle (e.g., Kimball et al., 2006; Zhang et al., 2008), indicate increasing trends in vegetation photosynthetic activity and net primary production in the Arctic over the past several decades." And with respect to what drove this welcome transformation of the tundra, Hudson and Henry say "it is likely that warming directly increased plant growth and reproduction and indirectly increased resource supply," while noting that "increased temperatures also lengthened the growing season, increased soil temperature, deepened the active [soil] layer, and consequently may have influenced nutrient uptake in the plant community."
But what if this area under study would have warmed even more? That was the focus of another paper by Hudson and Henry (2010) published the following year. In that study, the two authors used open-top chambers to passively warm an evergreen-shrub heath by 1.0-1.3°C over a period of fifteen years (during which time there was also a significant background warming) at the same location at Alexandra Fiord, Nunavut, Canada (79°N), which effort they describe as "the longest-running passive warming experiment in the Canadian Arctic." The results indicated, in the words of the two researchers, that "experimental warming did not strongly affect vascular plant cover, canopy height or species diversity, but it did increase bryophyte cover by 6.3% and decrease lichen cover by 3.5%," although they note that "temporal changes in plant cover were more frequent and of greater magnitude than changes due to experimental warming," as pointed out in their study published the year prior and highlighted above.
In another analysis, this time from Greenland, Madsen et al. (2011) compared the aboveground biomass of the graminoid marsh vegetation observed on Jameson Land, low Arctic, East Greenland in surveys conducted in 1982-1984, with similar surveys that were conducted in 2008, to see what had happened over the intervening period of significant global - and local - warming.
According to the five researchers, the data they obtained in 2008 yielded a standing crop biomass of 98.2 g/m2, which was 2.34 times greater than what had been measured in the same location in 1984. In further support of the validity of their observations, they note that "on Bylot Island, northeast Canada, graminoid above-ground production in wetlands has increased by 84% between 1990 and 2007, most likely as a consequence of climate warming," citing Cadieux et al. (2008), while adding that "on Svalbard, it is known that early snow melt has a dramatic positive effect on the density of nesting geese and their fecundity," citing Madsen et al. (2007) and noting that "the climate in East Greenland has been warming during the last 30 years." Thus, it would appear that many of Earth's higher-latitude terrestrial ecosystems might well be able to sustain considerably greater primary productivity, as well as much larger numbers of higher-trophic-level consumers, in a CO2-enriched and warmer world.
Introducing their study of the subject, Villarreal et al. (2012) write that "climate warming is pronounced at high northern latitudes (ACIA, 2005; Serreze, 2010)," and that "time series analysis of satellite remote sensing between 1982 and 2008 suggests that there has been a greening of arctic landscapes," citing Bhatt et al. (2010). More particularly, they note that "remotely detected changes in the normalized difference vegetation index (NDVI) of the arctic coastal plain near Barrow, Alaska, appear to be among the most dramatic recorded for much of the Arctic," once again citing Bhatt et al. (2010). However, they say "there is a general scarcity of ground-based studies that examine vegetation change in the Arctic over decade time scales," and it was this research niche that their study was designed to fill. More specifically, Villarreal et al. resampled (for species cover and presence, in 1999, 2008 and 2010) 330 marked plots at 33 sites that had been established in an area near Barrow at the northernmost point of the Alaskan Coastal Plain (71°18'N, 156°40'W) in 1972 as part of that year's International Biological Program.
Results indicated that over the 38-year study period, ecosystem diversity "increased for most plant communities, and wetter communities changed more than dry and moist plant communities." These findings are said by them to support other observational studies, such as that of Wilson and Nilsson (2009). And of the 19 species that comprised more than 80% of the overall relative cover, they say that five had higher relative cover in 2010 than 1972. In addition, they indicate that their results for changes in shrub cover "are somewhat consistent with other long-term observations and experimental studies that report increased shrub abundance (Myers-Smith et al., 2011; Tape et al., 2006; Walker et al., 2006; Sturm et al., 2001)," as well as warming-induced "treeline advancement (Lloyd, 2005; Danby and Hik, 2007)."
Also working in Alaska, Tape et al. (2006) analyzed repeat photography data from a photo study of the Colville River conducted between 1945 and 1953, as well as 202 new photos of the same sites that were obtained between 1999 and 2002, to determine the nature of shrub expansion in that region over the past half-century. This approach revealed, in their words, that "large shrubs have increased in size and abundance over the past 50 years, colonizing areas where previously there were no large shrubs." In addition, they say their review of plot and remote sensing studies confirms that "shrubs in Alaska have expanded their range and grown in size" and that "a population of smaller, intertussock shrubs not generally sampled by the repeat photography, is also expanding and growing." Taken together, they conclude that "these three lines of evidence allow us to infer a general increase in tundra shrubs across northern Alaska."
As for the cause of the shrub expansion and when it began, Tape et al. are inclined to attribute it to large-scale pan-Arctic warming; and from analyses of logistic growth curves, they estimate that the expansion began about 1900, "well before the current warming in Alaska (which started about 1970)." Hence, they conclude that "the expansion predates the most recent warming trend and is perhaps associated with the general warming since the Little Ice Age." These inferences appear reasonable, although the 80-ppm increase in the atmosphere's CO2 concentration since 1900 likely played a significant role in the shrub expansion as well.
In introducing their study, Elmendorf et al. (2012) state that "remote-sensing data indicate that contemporary climate warming has already resulted in increased productivity over much of the Arctic," noting that "normalized difference vegetation index (NDVI) values have increased over the tundra biome in recent years, indicating a greening of the tundra ecosystem coincident with climate warming trends," citing the studies of Pouliot et al. (2009) and Bhatt et al. (2010). However, they say that "plot-based evidence for vegetation transformation is not widespread," a deficiency which they proceed to correct by analyzing "change in tundra vegetation surveyed between 1980 and 2010 in 158 plant communities spread across 46 locations throughout the Arctic," the majority of which locations were from the continent of North America. And what did they find?
In the words of the 47 researchers hailing from 12 different countries who conducted the work, "we found biome-wide trends of increased height of the plant canopy and maximum observed plant height for most vascular growth forms; increased abundance of litter; increased abundance of evergreen, low-growing and tall shrubs; and decreased abundance of bare ground." These data confirm the findings of prior satellite assessments of the vegetative transformation of Earth's northernmost collection of landscapes over the past three decades, thanks not only to global warming, but also to the aerial-fertilization and water-use-efficiency-enhancing effects of atmospheric CO2 enrichment.
Further documentary evidence of enhanced plant productivity in the northern region of North America -- and the role of rising atmospheric CO2 in that enhancement -- comes from the study of Wang et al. (2006), who examined ring-width development in cohorts of young and old white spruce trees in a mixed grass-prairie ecosystem in southwestern Manitoba, Canada, where a 1997 wildfire killed most of the older trees growing in high-density spruce islands, but where younger trees slightly removed from the islands escaped the ravages of the flames. There, "within each of a total of 24 burned islands," in the words of the three researchers, "the largest dominant tree (dead) was cut down and a disc was then sampled from the stump height," while "adjacent to each sampled island, a smaller, younger tree (live) was also cut down, and a disc was sampled from the stump height."
After removing size-, age- and climate-related trends in radial growth from the ring-width histories of the trees, Wang et al. plotted the residuals as functions of time for the 30-year periods for which both the old and young trees would have been approximately the same age: 1900-1929 for the old trees and 1970-1999 for the young trees. During the first of these periods, the atmosphere's CO2 concentration averaged 299 ppm; during the second it averaged 346 ppm. Also, the mean rate-of-rise of the atmosphere's CO2 concentration was 0.37 ppm/year for first period and 1.43 ppm/year for the second.
The results of this exercise revealed that the slope of the linear regression describing the rate-of-growth of the ring-width residuals for the later period (when the air's CO2 concentration was 15% greater and its rate-of-rise 285% greater) was more than twice that of the linear regression describing the rate-of-growth of the ring-width residuals during the earlier period. As the researchers describe it, these results show that "at the same developmental stage, a greater growth response occurred in the late period when atmospheric CO2 concentration and the rate of atmospheric CO2 increase were both relatively high," and they say that "these results are consistent with expectations for CO2-fertilization effects." In fact, they say "the response of the studied young trees can be taken as strong circumstantial evidence for the atmospheric CO2-fertilization effect."
Another thing Wang et al. learned was that "postdrought growth response was much stronger for young trees (1970-1999) compared with old trees at the same development stage (1900-1929)," and they add that "higher atmospheric CO2 concentration in the period from 1970-1999 may have helped white spruce recover from severe drought." In a similar vein, they also determined that young trees showed a weaker relationship to precipitation than did old trees, noting that "more CO2 would lead to greater water-use efficiency, which may be dampening the precipitation signal in young trees."
Taken together, the results presented here for the high latitude region of North America paint a picture of the planet's terrestrial vegetation that is opposite of that promulgated by the world's climate alarmists. Instead of heading down the road of environmental degradation toward extinction, land-based plants of the Arctic and near-Arctic regions of North America are thriving, thanks in large part to the ongoing rise in the atmosphere's CO2 concentration and global warming.