




The State of Earth's Terrestrial Biosphere:
How is it Responding to Rising Atmospheric CO2 and Warmer Temperatures?
Continental-Scale Analyses of Terrestrial Productivity: Europe
Turing to Europe, using an empirically-based mechanistic model of Mediterranean shrub vegetation, Osborne et al. (2000) set out to address two important questions: (1) Has recent climate change, especially increased drought, negatively impacted Mediterranean shrublands? and (2) Has the historical increase in the air's CO2 concentration modified this impact? The data-based model they employed suggests that the warming and reduced precipitation experienced in the Mediterranean area over the past century should have had negative impacts on net primary production and leaf area index. When the measured increase in atmospheric CO2 concentration experienced over the period was factored into the calculation, however, these negative influences were overpowered, with the net effect that both measures of vegetative prowess increased: net primary productivity by 25% and leaf area index by 7%. These results, in their words, "indicate that the recent rise in atmospheric CO2 may already have had significant impacts on productivity, structure and water relations of sclerophyllous shrub vegetation, which tended to offset the detrimental effects of climate change in the region."
With respect to model-predicted changes in Earth's precipitation regime, a doubling of the air's CO2 content is projected to lead to a modest intensification of the planet's hydrologic cycle. In the case of the Mediterranean region over the last century, however, there has been a recent tendency toward drier conditions. Hence, the specific case investigated by Osborne et al. represents a much-worse-case scenario than what is predicted by current climate models for the Earth as a whole. Nevertheless, the area's vegetation has done even better than it did before the climatic change, thanks to the over-powering beneficial biological effects of the concurrent rise in the air's CO2 content.
In another model-based study, Cheddadi et al. (2001) employed a standard biogeochemical model (BIOME3) - which uses monthly temperature and precipitation data, certain soil characteristics, cloudiness and atmospheric CO2 concentration as inputs - to simulate the responses of various biomes in the region surrounding the Mediterranean Sea to changes in both climate (temperature and precipitation) and the air's CO2 content. Their first step in this endeavor was to validate the model for two test periods: the present and 6000 years before present (BP). Recent instrumental records provided actual atmospheric CO2, temperature and precipitation data for the present period; while pollen data were used to reconstruct monthly temperature and precipitation values for 6000 years BP, and ice core records were used to determine the atmospheric CO2 concentration of that earlier epoch. These efforts suggested that winter temperatures 6000 years ago were about 2°C cooler than they are now, that annual rainfall was approximately 200 mm less than today, and that the air's CO2 concentration averaged 280 ppm, which is considerably less than the value of 345 ppm the researchers used to represent the present, i.e., the mid-point of the period used for calculating 30-year climate normals at the time they wrote their paper. Applying the model to these two sets of conditions, they demonstrated that "BIOME3 can be used to simulate ... the vegetation distribution under ... different climate and CO2 conditions than today."
Cheddadi et al.'s next step was to use their validated model to explore the vegetative consequences of an increase in anthropogenic CO2 emissions that pushes the air's CO2 concentration to a value of 500 ppm and its mean annual temperature to a value 2°C higher than today's mean value. The basic response of the vegetation to this change in environmental conditions was "a substantial southward shift of Mediterranean vegetation and a spread of evergreen and conifer forests in the northern Mediterranean."
More specifically, in the words of the researchers, "when precipitation is maintained at its present-day level, an evergreen forest spreads in the eastern Mediterranean and a conifer forest in Turkey." Current xerophytic woodlands in this scenario become "restricted to southern Spain and southern Italy and they no longer occur in southern France." In northwest Africa, on the other hand, "Mediterranean xerophytic vegetation occupies a more extensive territory than today and the arid steppe/desert boundary shifts southward," as each vegetation zone becomes significantly more verdant than it is currently.
What is the basis for these positive developments? Cheddadi et al. say "the replacement of xerophytic woodlands by evergreen and conifer forests could be explained by the enhancement of photosynthesis due to the increase of CO2." Likewise, they note that "under a high CO2 stomata will be much less open which will lead to a reduced evapotranspiration and lower water loss, both for C3 and C4 plants," adding that "such mechanisms may help plants to resist long-lasting drought periods that characterize the Mediterranean climate."
Contrary to what is often predicted for much of the world's moisture-challenged lands, therefore, the authors were able to report that "an increase of CO2, jointly with an increase of ca. 2°C in annual temperature would not lead to desertification on any part of the Mediterranean unless annual precipitation decreased drastically," where they define a drastic decrease as a decline of 30% or more. Equally important in this context is the fact that Hennessy et al. (1997) have indicated that a doubling of the air's CO2 content would in all likelihood lead to a 5 to 10% increase in annual precipitation at Mediterranean latitudes, which is also what is predicted for most of the rest of the world. Hence, the results of the present study - where precipitation was held constant - may validly be considered to be a worst-case scenario, with the true vegetative response being even better than the good-news results reported by Cheddadi et al., even when utilizing what we believe to be erroneously-inflated global warming predictions.
Introducing their work, Bellassen et al. (2011) write that "several parties to the United Nations Framework Convention on Climate Change (UNFCCC) are calling for 'forward-looking baselines'," so that country-specific scenarios based on forest age structure could be used "to credit only the part of the forest sink going beyond business-as-usual practices." And they thus proceed to derive such a baseline for all of Europe.
Using ORCHIDEE-FM - a process-based vegetation model that differs from earlier versions of ORCHIDEE by "its explicit representation of stand growth and idealized forest management" - Bellassen et al. applied the model on a grid across Europe to "simulate changes in the net ecosystem productivity (NEP) of forests with and without changes in climate, CO2 and age structure." In doing so, the six scientists report that the model they used "simulates carbon stocks and volume increments that are comparable with inventory-derived estimates at country level for 20 European countries," providing "an upwards trend of forest NEP of 1 ± 0.5 g C/m2/year between 1950 and 2000 across the EU 25," ending with "a mean European forest NEP of 175 ± 52 g C/m2/year in the 1990s" (see Figure 15). And they say that "61% of the change in NEP [over the last half of the 20th century] was attributed to changes in CO2, 26% to changes in climate, and 13% to changes in forest age structure."
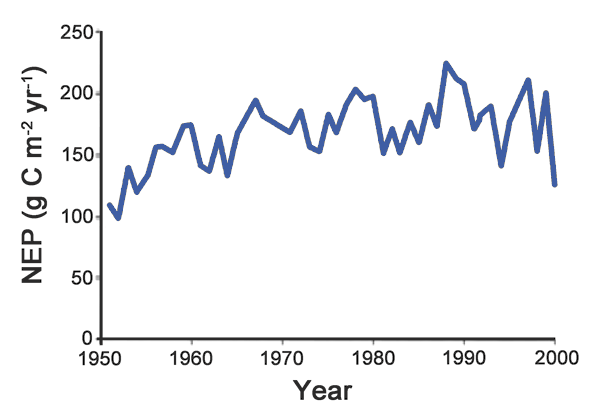
Figure 15. Simulated reconstruction of Net Ecosystem Productivity (NEP) in European forests from 1951-2000. Adapted from Bellassen et al. (2011).
As intriguing as these model-based studies are, however, it is important to examine this issue through the lens of real-world data to see how plant productivity has responded to modern warming, which the IPCC claims is unprecedented over the past one to two thousand years. And, fortunately, a number of scientists have done just that.
Allen et al. (1999), for example, analyzed sediment cores from a lake in southern Italy and from the Mediterranean Sea, developing high-resolution climate and vegetation data sets for this region over the last 102,000 years. These materials indicated that rapid changes in vegetation were well correlated with rapid changes in climate, such that complete shifts in natural ecosystems would sometimes occur over periods of less than 200 years. Over the warmest portion of the record (the Holocene), the total organic carbon content of the vegetation reached its highest level, more than doubling values experienced over the rest of the record, while other proxy indicators revealed that during the more productive woody-plant period of the Holocene, the increased vegetative cover also led to less soil erosion. The results of this study thus demonstrate that the biosphere can successfully respond to rapid changes in climate. As the 15 researchers involved in the work put it, "the biosphere was a full participant in these rapid fluctuations, contrary to widely held views that vegetation is unable to change with such rapidity." Furthermore, their work revealed that warmer was always better in terms of plant growth; and, therefore, it is likely that future warming in this region may return it to a higher level of biological productivity than what it currently exhibits.
Examining a much shorter time period was Bert et al. (1997), who calculated a 120-year (1860-1980) history of intrinsic water-use efficiency (defined as the ratio of CO2 assimilation rate to stomatal conductance for water vapor) for silver fir (Abies alba Mill.) trees, based on δ13C data obtained from cores extracted from individual trees of this species that grew within 208 pure stands in the Jura Mountains near the border that separates France and Switzerland. Results indicated that from 1860 to 1930 there was little net change in silver fir water-use efficiency; but over the next half-century (1930 to 1980), when the atmosphere's CO2 concentration rose at a rate that was more than three times greater than its rate-of-rise over the earlier period, this important tree physiological property rose by approximately 30% (see Figure 16). With regard to their findings, the three researchers state that their results - which were "obtained at the level of mature trees" - are "consistent with the physiological effects of increasing CO2 concentrations as observed in controlled experiments on young seedlings," and that they are additionally "consistent with the strong increases in radial growth observed for Abies alba in western Europe over the past decades."
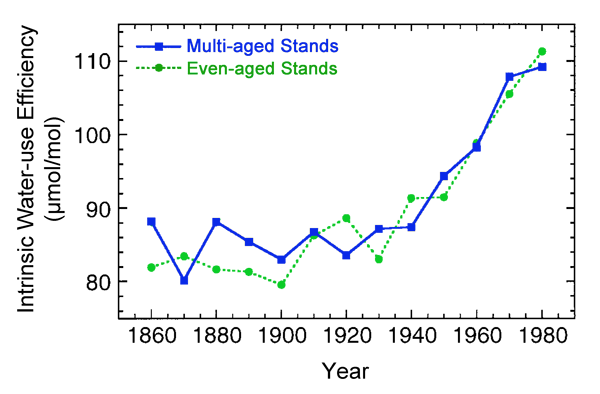
Figure 16. Intrinsic water-use efficiency (the ratio of CO2 assimilation rate to stomatal conductance for water vapor) calculated from tree rings sampled in multi-aged and even-aged forest stands from the Jura Mountains in Eastern France for the period 1860-1980. Adapted from Bert et al. (1997).
Working in the Vienna basin of Austria in the European Eastern Alps, Leal et al. (2008) discovered what they describe as "a very clear change in the sensitivity of the growth rate of [black pine (Pinus nigra)] tree stems to water availability in the late 20th century," noting that "trees previously sensitive to spring-summer drought show a lack of response to this climatic parameter in recent decades." That is to say, as they explain it, that "tree-ring indices were larger in the second half of the 20th century than predicted given prevailing spring-summer drought conditions and the previous sensitivity of growth to these conditions." In addition, they found "a decrease in correspondence between the occurrence of extreme events in precipitation and rate of change of growth," such that "in the second half of the century this correspondence was not significant," and that "recent extreme droughts did not result in the formation of very narrow rings, which means the droughts were not as limiting to tree growth as they had been in the past."
The five researchers concluded their paper by suggesting that the greater atmospheric CO2 concentrations of the latter decades of the 20th century "induced improved water-use efficiency enabling P. nigra growing in the Vienna basin to avoid the impact of recurrent dry conditions," which phenomenon has also been observed in many other parts of the world in a number of different tree species, which is but another indication of the propensity of the ongoing rise in the air's CO2 content to promote a greening of the Earth.
Martinez-Vilalta et al. (2008) used tree-ring data from the Catalan Ecological and Forest Inventory "to study the temporal variability of Scots pine stem radial growth (period 1901-1997) across a relatively large region (Catalonia, NE Spain) situated close to the southern limit of the distribution of the species." This inventory "included a total of 10,664 plots randomly distributed throughout the forested area of Catalonia." Scots pine was present in 30.2% of the plots, and it was the dominant tree species in 18.4% of them.
In describing their findings, the researchers say they found "an overall increase of 84% in Scots pine BAI [basal area increment] during the 20th century (see Figure 17), consistent with most previous studies for temperate forests" and in harmony with the greening of the Earth phenomenon that has accompanied the historical increase in the air's CO2 content. And in this regard, they make a point of stating that "this trend was associated with increased atmospheric CO2 concentration," which they interpreted to be "a fertilization effect." What is more, the five scientists also note that over the same time period there was "a marked increase in temperature across the study region (0.19°C per decade on average)," and they report that "this warming had a negative impact on radial growth, particularly at the drier sites." However, they found that "its magnitude was not enough to counteract the fertilization effect."
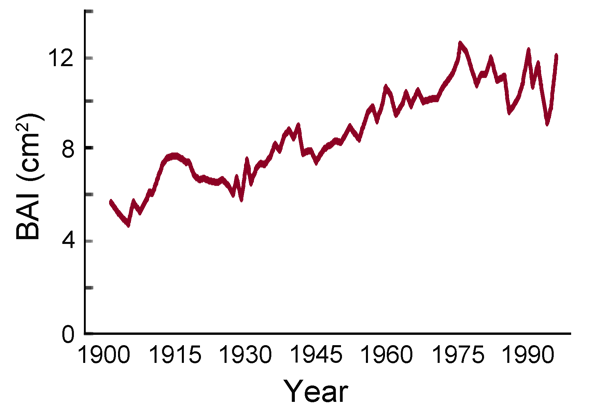
Figure 17. Basal area increment (BAI) for Scots pine trees growing in NE Spain over the period 1901-1997. Adapted from Martinez-Vilalta et al. (2008).
Noting that "protected areas provide excellent opportunities for [determining] baseline descriptions and trends that ... can be used to evaluate the impact of global environmental change on terrestrial ecosystem functioning," Alcaraz-Segura et al. (2008) employed satellite-derived normalized difference vegetation index (NDVI) data - which provide a measure of net primary production that is described by them as "the most integrative indicator of ecosystem functioning" - to "evaluate the impact of global environmental change on terrestrial ecosystem functioning of [Spain's] national parks," which provides a sound basis for determining what could have been expected to have occurred throughout the rest of the country and much of Europe (Julien et al., 2006) independent of confounding effects not related to global environmental change. So what did they find?
The four researchers, hailing from Argentina, Spain and the United States, report that "most parks showed areas with positive NDVI trends that tended to have higher proportions of Mediterranean coniferous and mixed forests, oro-Mediterranean scrublands, heathlands, maquis and garrigues," while "negative trends were scarce." Alcaraz-Segura et al. conclude the report of their findings by stating that "protected areas are changing in the short term and, at least in terms of vegetation greenness, they are changing in a directional way," such that "a large part of the Spanish National Parks is intercepting more photosynthetically active radiation than in the past."
Working at a site just three kilometers from the Abisko Scientific Research Station (68°21'N, 18°49'E) in the Northern Swedish Scandes, Hallinger et al. (2010) studied male plants of the medium-sized Juniperus nana shrub, collecting the main stems of five to eight shrubs every hundred meters of elevation until the shrub zone ended. Ring-width measurements on these stems were then performed, as they describe it, "to measure radial and vertical growth, to track growth changes over time, to age the shrub individuals and to correlate annual shrub growth with climate," the characteristics of which latter factor were derived from records of the nearby Abisko Station. And what did their measurements reveal?
According to the three researchers, their analysis "documented a distinct increase in radial and vertical growth rates of J. nana shrubs during recent decades in the subalpine zone of North Sweden" (see Figure 18), and they say that "the age structure of shrubs along the elevational gradient provides evidence that an upslope advance of the altitudinal shrubline is underway." In addition, they state that they "observed significant, strong and stable correlations between annual ring width and summer temperatures (June, July, August)," and that "the acceleration of radial and vertical growth since 1970 also coincides with the recent three decades of rising arctic air temperatures and the warming trend of 0.2°C per decade for the average temperature since 1956 at Abisko." These findings, in their words, add to the "mounting evidence that shrubs are expanding into alpine and arctic areas because of climate warming," and they note that "this expansion occurs in both evergreen and deciduous shrub types," citing the additional findings of Forbes et al. (2009).
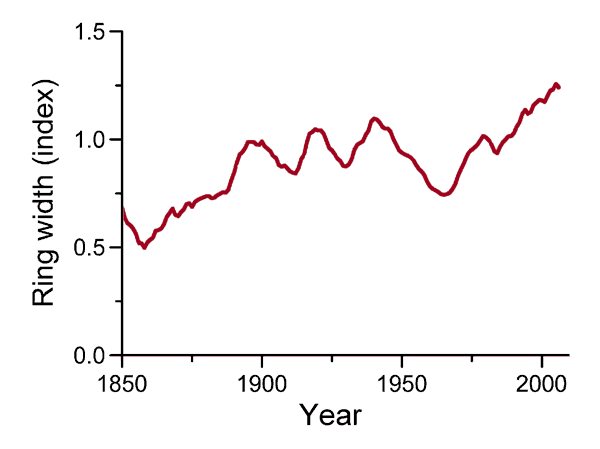
Figure 18. Indexed radial growth of juniper trees in the Northern Swedish Scandes over the period 1850-2006, smoothed with a 10-year filter. Adapted from Hallinger et al. (2010).
Also working in the Swedish Scandes, Kullman (2010a) presented what he called "an integrative review of results from long-term monitoring of subalpine/alpine vegetation," from which he derived "tentative projections of landscape transformations in a potentially warmer future," which were based on "actual observations and paleoecological data (Kullman and Kjallgren, 2006; Kullman 2006)." In doing so, the professor of physical geography at Sweden's Umea University indicates that post-Little Ice Age warming has, at long last, broken the back of "a multi-millennial trend of plant cover retrogression" and "floristic and faunal impoverishment, all imposed by progressive and deterministic neoglacial climate cooling," as he reports that the "upper range margin rise of trees and low-altitude (boreal) plant species, expansion of alpine grasslands and dwarf-shrub heaths are the modal biotic adjustments during the past few decades, after a century of substantial climate warming." Currently, therefore, the situation is one where "alpine plant life is proliferating, biodiversity is on the rise and the mountain world appears more productive and inviting than ever." And he makes it very clear that "in contrast to model predictions, no single alpine plant species has become extinct, neither in Scandinavia nor in any other part of the world in response to climate warming over the past century," citing, in addition to his own studies, the work of Pauli et al. (2001, 2007), Theurillat and Guisan (2001), and Birks (2008).
So just how great is the recent proliferation of plant life of which Kullman writes? In a contemporaneous study published in the Nordic Journal of Botany, he provides an answer (Kullman, 2010c), reporting what he learned from species inventories he conducted on the uppermost 20 meters of four high-mountain summits in the Swedish Scandes (Kullman 2007a,b), the results of which he compared with the findings of "historical species inventories from the early 1950s, executed by a highly competent and experienced botanist (Kilander, 1955)," which endeavor, in his words, "can be seen as an evaluation of a full-scale 'natural experiment' (cf. Grabherr et al., 2001)."
Specifically, Kullman (2010c) writes that the species pools at the tops of the studied mountains have (1) "increased by 60-170% since the 1950s," that (2) "some of the invading species are new to the alpine tundra, with more silvine and thermophilic properties than the extant alpine flora," and -- last of all, that (3) "not a single species of the original flora has disappeared from any of the summits." Concluding, Kullman thus writes that "the alpine flora appears to be more adaptive and responsive to climate change than generally believed," and that "overall, a richer, greener and more productive alpine world has emerged in the wake of the recent climate warming episode (Kullman, 2010a, 2010b)."
Still in Sweden, but working on an east-facing slope of the Slattatjakka/Njulla mountains (68°21'N, 18°49'W) in the Abisko Valley about 200 km north of the Arctic Circle, Rundqvist et al. (2011) write that they documented "rapid and substantial increases in the abundance of prominent tree and shrub species near [the] tree-line and forest-line in sub-Arctic Sweden," and that they "recorded an invasion by a thermophilic tree species not present in the plots 34 years ago."
In discussing their findings, the seven Swedish scientists describe how they mesh well with those of many other researchers, noting that "there is an indication that the shrub layer near the tree-line has expanded, since the 1930s, in the Abisko area (Enquist et al., 1933; Sandberg, 1963)," while "data from Canada, Fennoscandia, Alaska and Russia reveal that there is a Pan-Arctic expansion of shrubs and trees in progress (e.g. Kullman, 2002; Tommervik et al., 2004; ACIA, 2005; Tape et al., 2006; Karlsson et al., 2007; Olofsson et al., 2009; Hallinger et al., 2010; Hendenas et al., 2011)." And they state that the change in shrubs and small trees they observed is "consistent with anticipated changes due to climate change and reduced herbivory," which change in climate, in their words, "could be interpreted as an ongoing natural re-establishment of plants at higher altitudes due to a natural increase in the temperature since the 'Little Ice Age' (Kammer et al., 2007)."
Writing as background for their study, Hedenas et al. (2011) state that "during the last 15 years, there has been an increasing focus on how climate change has and will affect the distribution and extent of ecosystems around the globe including alpine and Arctic areas (e.g., Callaghan et al., 2005)," and in this regard they report that "field studies and remote sensing have revealed a recent increase in altitude of the tree line (e.g., Kullman, 2002)," as well as "an extension and increased cover of mountain birch forest (Tommervik et al., 2009; Rundqvist et al., 2011)." More specifically, they say that Tommervik et al. have determined that "tree biomass has doubled over a 43-year period, within an area of Finnmarksvidda, and Rundqvist et al. have observed an increased density and cover of mountain birch in the treeline over the last three decades, within an area near Abisko village."
In a continuation of these types of studies, Hedenas et al. say that in 2010 they re-surveyed shrub, tree and vegetation data at 549 plots grouped into 61 clusters that were originally surveyed in 1997 in two areas close to the Abisko village, which is located approximately 200 km north of the Arctic Circle at 68°20'N, 18°50'E. Their results indicated that "tree basal area and biomass increased by 19% between 1997 and 2010 with the main increase occurring in established birch forest" (see Figure 19), and they say that this result "concurs with the results of other studies which suggest that there has been a general increase in cover and biomass of trees and shrubs in sub-Arctic and Arctic areas," additionally citing in this regard, the studies of Sturm et al. (2001), Tape et al. (2006), Danby and Hik (2007), Forbes et al. (2010a), Hallinger et al. (2010) and Van Bogaert et al. (2011). What is more, the 19% net increase in biomass occurred in spite of the increased browsing pressure provided by an increasing reindeer population over the period of their study, as well as periodic outbursts of geometrid moths - which severely defoliated the birch trees in their study area in 2004 (Babst et al., 2010). As for the cause of this welcome phenomenon, they say "it has been suggested that increased nutrient availability associated with higher soil temperatures, and a longer growing season could underpin increased tree and shrub abundance and biomass in the Arctic (e.g., Chapin, 1983; Weih and Karlsson, 1997; Hartley et al., 1999)," as a result of "a delayed re-expansion of shrubs and trees following the 'Little Ice Age'," as suggested by Grubb (2008).

Figure 19. Change in mean tree biomass by altitude over the period 1997-2010 from various locations throughout the Swedish sub-Arctic. Colors reveal the direction of the change (green = biomass increase, yellow = no change, and red = biomass decrease), while arrows indicate the magnitude of the change over the 13-year period. Adapted from Hedenas et al. (2010).
Noting that forests are exposed to a changing environment and that "responses to recent climate change start to become visible if observation periods become long enough," Pilegaard et al. (2011) present the results of continuous CO2 flux measurements that they made above a mature Danish beech stand in the "Lille Bogeskov" forest located near Soro on the island of Zealand (55°29'13"N, 11°38'45"E) over the period 1996-2009, describing the long-term changes they observed and relating them to possible causes.
According to the researchers, they observed "significant linear trends of increasing gross ecosystem exchange (GEE: 29 g C/m2/year) and increasing net ecosystem exchange (NEE: 23 g C/m2/year), while the positive trend for ecosystem respiration (RE: 5 g C/m2/year) was not significant." They also state that "the length of the carbon uptake period increased by 1.9 day/year, whereas there was a non-significant increase of 0.3 day/year in the leafed period," which means, in their words, that "the leaves stay active longer." Nevertheless, they indicate that "the increase in the carbon uptake period explained only part of the increasing NEE (9 gC/m2/year)." And noting that "the maximum rate of photosynthetic assimilation increased by 15% during the 14-year period," they speculate that the increase in canopy carbon uptake capacity "could be due to a combination of [the] increase in atmospheric CO2, higher summer precipitation, and increased availability of nitrogen."
Working in the Komi Republic in the northeast European sector of Russia, Lopatin et al. (2006) (1) collected discs and cores from 151 Siberian spruce trees and 110 Scots pines from which they developed ring-width chronologies that revealed yearly changes in forest productivity, (2) developed satellite-based time series of NDVI for the months of June, July, August over the period 1982-2001, (3) correlated their site-specific ring-width-derived productivity histories with same-site NDVI time series, (4) used the resulting relationship to establish six regional forest productivity histories for the period 1982-2001, and (5) compared the six regional productivity trends over this period with corresponding-region temperature and precipitation trends. For all six vegetation zones of the Komi Republic, this work indicated that the 1982-2001 trends of integrated NDVI values from June to August were positive, and that the "increase in productivity reflected in [the] NDVI data [was] maximal on the sites with increased temperature and decreased precipitation."
In discussing their findings, the three scientists state that "several studies (Riebsame et al., 1994; Myneni et al., 1998; Vicente-Serrano et al., 2004) have shown a recent increase in vegetation cover in different world ecosystems." What is special about their study, as they describe it, is that "in Europe, most forests are managed, except for those in northwestern Russia [the location of their work], where old-growth natural forests are dominant (Aksenov et al., 2002)." Consequently, and because of their positive findings, they say we can now conclude that "productivity during recent decades also increased in relatively untouched forests," where non-management-related "climate change with lengthening growing season, increasing CO2 and nitrogen deposition" are the primary determinants of changes in forest productivity.
In one final study that integrates all of Europe, Julien et al. (2006) "used land surface temperature (LST) algorithms and NDVI [Normalized Difference Vegetation Index] values to estimate changes in vegetation in the European continent between 1982 and 1999 from the Pathfinder AVHRR [Advanced Very High Resolution Radiometer] Land (PAL) dataset." This program revealed that arid and semi-arid areas (Northern Africa, Southern Spain and the Middle East) have seen their mean LST increase and NDVI decrease, while temperate areas (Western and Central Europe) have suffered a slight decrease in LST but a more substantial increase in NDVI, especially in Germany, the Czech Republic, Poland and Belarus. In addition, parts of continental and Northern Europe have experienced either slight increases or decreases in NDVI while LST values have decreased. Considering the results in their totality, the Dutch and Spanish researchers concluded that, over the last two decades of the 20th century, "Europe as a whole has a tendency to greening," and much of it is "seeing an increase in its wood land proportion."