




Volume 16, Number 23: 5 June 2013
In a significant new study published in Global Change Biology, Koutavas (2013) describes how he employed dendrochronology (the study of tree growth rings) to investigate potential growth-climate relationships in Greek fir trees growing on Ainos Mountain on the island of Cephalonia in the Ionian Sea, which region, in his words, "has recently experienced some of the largest climatic drying in the globe." And as a result of what he calls this "pronounced shift to greater aridity in recent decades," he states that "this region offers unique opportunities to investigate [tree] growth responses to drought, particularly with relevance to elevated atmospheric CO2, which is thought to confer moisture conservation benefits in arid and semiarid settings by allowing plants to reduce their stomatal conductance and increase their water utilization efficiency (WUE)," citing the work of Wullschleger et al. (2002), Morgan et al. (2004) and Huang et al. (2007).
Setting out to do just that, Koutavas developed growth indices from cores he extracted from 23 living Greek fir (Aibes cephalonica) trees located between elevations of 1300 and 1600 meters for the period AD 1820-2007; and from those indices he reports finding that the growth of the trees historically has been "limited by growing-season moisture in late spring/early summer, most critically during June." However, he further finds that "this simple picture obscures a pattern of sharply evolving growth-climate relationships during the 20th century," wherein the "correlations between growth and June temperature, precipitation and Palmer Drought Severity Index were significantly greater in the early 20th century but later degraded and disappeared" (see Figure 1 below). In fact, Koutavas writes that "by the late 20th-early 21st century, there remains no statistically significant relationship between moisture and growth."
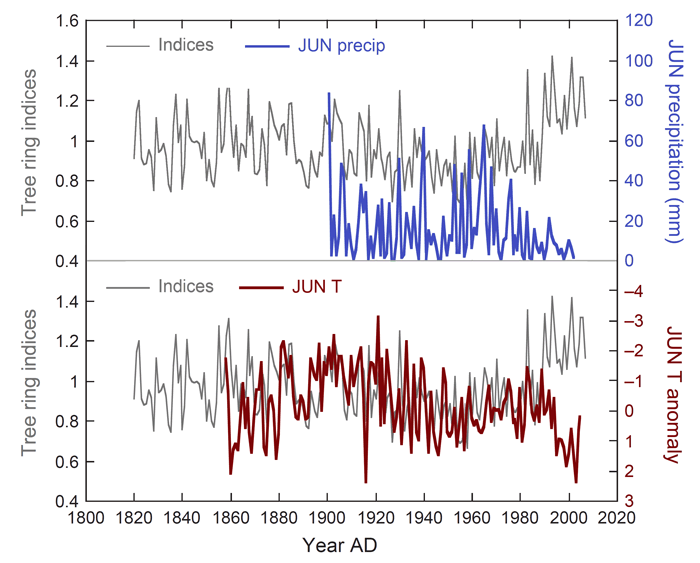
Figure 1. Greek fir tree ring indices plotted with June precipitation and temperature anomalies, illustrating the diverging trends among the data sets in the latter portion of the records. Adapted from Koutavas (2013).
But the story does not end there. According to Koutavas, in spite of the "pronounced shift to greater aridity in recent decades," tree growth in the region actually experienced "a net increase over the last half-century, culminating with a sharp spike in AD 1988-1990," which growth increase implies that the trees have acquired a "markedly enhanced resistance to drought." So how did that happen?
Such observations, in the words of Koutavas "are most consistent with a significant CO2 fertilization effect operating through restricted stomatal conductance and improved water-use efficiency." And he also opines that "if this interpretation is correct" - and what other interpretation could there possibly be? - "atmospheric CO2 is now overcompensating for growth declines anticipated from drier climate, suggesting its effect is unusually strong and likely to be detectable in other up-to-date tree-ring chronologies from the Mediterranean."
There we have it. The increasing concentration of atmospheric CO2, as illustrated in the figure below, appears to be the most important factor driving recently enhanced growth rates of Greek fir trees. And it is doing so in spite of unfavorable moisture conditions and declining temperatures that should be causing growth declines. Not bad for a growth-promoting and life-sustaining molecule that some have incorrectly labeled a "pollutant."
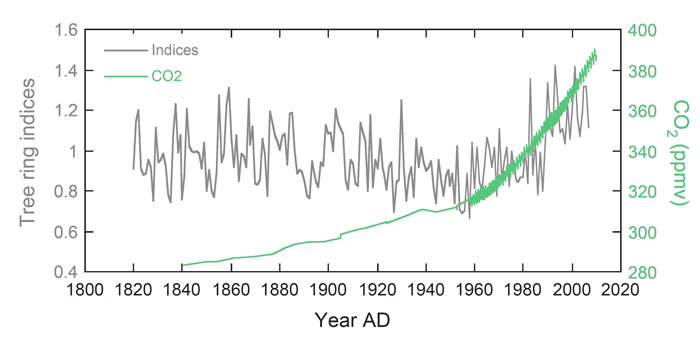
Figure 2. Greek fir tree ring indices plotted with historic atmospheric CO2 concentration, illustrating the high degree of correlation that exists among the two variables since the mid-20th century. Adapted from Koutavas (2013).
Sherwood, Keith and Craig Idso
References
Huang, J.-G., Bergeron, Y., Denneler, B., Berninger, F. and Tardiff, J. 2007. Response of forest trees to increased atmospheric CO2. Critical Reviews in Plant Science 26: 265-283.
Koutavas, A. 2013. CO2 fertilization and enhanced drought resistance in Greek firs from Cephalonia Island, Greece. Global Change Biology 19: 529-539.
Morgan, J.A., Pataki, D.E., Korner, C., Clark, H., Del Grosso, S.J., Grunzweig, J.M., Knapp, A.K., Mosier, A.R., Newton, P.C., Kiklaus, P.A., Nippert, J.B., Nowak, R.S., Parton, W.J., Polley, H.W. and Shaw, M.R. 2004. Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 140: 11-25.
Wullschleger, S.D., Tschaplinski, T.J. and Norby, R.J. 2002. Plant water relations at elevated CO2 - implications for water-limited environments. Plant, Cell and Environment 25: 319-331.