




Separating Scientific Fact from Personal Opinion
A critique of the 26 April 2007 testimony of James E. Hansen made to
the Select Committee of Energy Independence and Global Warming
of the United States House of Representatives entitled
"Dangerous Human-Made Interference with Climate"
Prepared by
Sherwood B. Idso and Craig D. Idso
Center for the Study of Carbon Dioxide and Global Change
6 June 2007
If there is any human enterprise that should be free of appeal to authority, it is science, where observation and impartial analysis are supposed to reign supreme. However, when the outcome of an ongoing scientific investigation is perceived to be a powerful catalyst for governmental action by the world's community of nations, and when the leading policy prescription for those actions is something akin to a massive restructuring of the way the energy that runs the modern world is produced, distributed and used - and especially if the policy is developed before all pertinent data have been acquired and properly analyzed - this principle can easily be forgotten. In such circumstances, and even more so if the subject being studied is extremely complex - such as how human activity will impact global climate centuries into the future - and when a divergence of views develops because of ambiguities in the observations and different methods of analysis, it is important that personal opinion be clearly differentiated from demonstrable fact. Sadly, however, this distinction is hard to make on a consistent basis, even for some of the very best of the world's scientists.
A case in point is the testimony that was presented by Dr. James E. Hansen to the Select Committee of Energy Independence and Global Warming of the United States House of Representatives on 26 April 2007. Being a recognized expert in the field of climatology, particularly climate modeling, and being considered by many to be perhaps the world's foremost authority on the "greenhouse effect" of anthropogenic CO2 emissions, Hansen's statements are typically regarded as expressions of fact. In many cases, however, they are merely his opinions. Hence, it is important to compare what Hansen says in his testimony with what has been learned through the practice of science by a large number of other researchers, whose vast arrays of observations and detailed analyses sometimes lead to conclusions that differ significantly from what Hansen contends.
In the materials that follow, we present such a comparison, focusing on a number of key subjects addressed by Hansen. These topics include: (1) ice sheet disintegration, (2) sea level trends, (3) atmospheric methane concentrations, (4) climates of the past, (5) predicted warming-induced extinctions of terrestrial plants and animals, (6) the CO2-induced preservation of terrestrial species, and (7) predicted CO2-induced extinctions of calcifying marine organisms. In addition, we discuss a number of other topics that Hansen addresses in less detail, including: (1) positive vs. negative climate feedbacks, (2) effects of drought on agriculture in a CO2-enriched world, (3) sea level rise over the next hundred years, (4) the adaptability of living organisms to rising sea levels, (5) the "dangerous" level of atmospheric CO2, (6) the magnitude of climate forcing due to a doubling of the air's CO2 content, (7) empirical evaluations of earth's climate sensitivity, (8-10) the ability of man to control global climate, (11-14) the need to act now to reduce CO2 emissions, and (15) the role of morality in the debate over what to do, or not do, about anthropogenic CO2 emissions. And when Hansen's testimony is compared with what has been revealed by the scientific investigations of a diverse assemblage of highly competent researchers in a wide variety of academic disciplines, we find that he paints a very different picture of the role of anthropogenic CO2 emissions in shaping the future fortunes of man and nature alike than what is suggested by that larger body of work.
Hansen's testimony is divided into five parts: (1) Summary, (2) Basis for Testimony, (3) Crystallizing Science, (4) Metrics for Dangerous Climate Change, and (5) Four-Point Strategy to Stabilize Climate. We will begin our critique of the document with a brief analysis of what Hansen says is its foundation, i.e., his Section 2: Basis for Testimony.
Six papers in various stages of preparation for publication in peer-reviewed scientific journals form the basis for Hansen's testimony. The first, written by Hansen and 46 co-authors, is entitled "Dangerous human-made interference with climate: A GISS modelE study" and is listed as being "in press" in Atmospheric Chemistry and Physics. The second paper, written by Hansen and five co-authors, is entitled "Climate change and trace gases" and is listed as being "in press" in the Philosophical Transactions of the Royal Society A. The third paper, also written by Hansen and 46 co-authors, is entitled "Climate simulations for 1880-2003 with GISS modelE" and is listed as being "in press" in Climate Dynamics. The fourth paper, written by Hansen alone, is entitled "Scientific reticence and sea level rise" and is listed as being "accepted for publication" by Environmental Research Letters. The fifth paper, again by Hansen alone, is entitled "State of the wild: Perspective of a climatologist" and is listed as being "accepted" by an unnamed journal. The sixth paper, where Hansen appears as the second of two authors, is entitled "Implications of 'peak oil' for atmospheric CO2 and climate" and is listed as being a "first draft" prepared for Geophysical Research Letters.
In perusing these manuscripts, it is readily apparent they either deal with, or are based upon, scenario-driven climate-model projections, which obviously can be no better than the physics, chemistry and biology upon which they are based, as well as the scenarios that drive them. To be of any prognostic value, therefore, the models must include, and correctly characterize, all of the physical, chemical and biological phenomena that significantly impact the planet's climate, which is something most climate modelers would probably admit they have not yet achieved to the degree they would like. But are they close enough? Our only way of answering this question is to see if what the models portend about the future compares favorably with what they suggest about the past. Of course, the models could accidentally give the "right answers," but there is no other course of action we can take at the present time; and, hence, this is what we will do in evaluating Hansen's testimony, for if the models give the wrong answers about the recent past, we can be confident they are not up to the task of correctly inferring the future.
The core concept of Hansen's testimony is that the earth "is close to dangerous climate change, to tipping points of the system with the potential for irreversible deleterious effects." However, this contention, like the many other claims Hansen makes, is neither a self-evident verity nor a proven fact. It is merely an opinion. And to raise it to a loftier status requires that there be real-world evidence for the changes the climate models suggest should occur in response to increasing atmospheric CO2 concentrations and rising air temperatures. This requirement is all the more justified in light of the fact that air temperatures of the last quarter-century are typically claimed by climate alarmists to have been unprecedented for at least the past two thousand years (Mann and Jones, 2003; Mann et al., 2003) - and possibly for close to a million years, if one believes Hansen et al. (2006) - while the atmosphere's current CO2 concentration is greater than it may have been for tens of millions of years (Pagani et al., 1999).
So what are the major climate changes and associated catastrophic consequences that are suggested by the climate models? And are there any signs they may already be in process of developing? The "sharpest criterion" for defining dangerous climate change, in the words of Hansen, "is probably maintenance of long-term sea level close to the present level," and in this regard he says that "sea level is already rising at a rate of 3.5 cm per decade and the rate is accelerating [our italics]," due, he would have us believe, to "ice sheet disintegration." But are there any real-world data to support this claim?
Ice Sheet DisintegrationA good perspective on this issue is provided in the 16 March 2007 issue of Science by Shepherd and Wingham (2007), who review what is known about sea-level contributions arising from wastage of the Antarctic and Greenland Ice Sheets, focusing on the results of 14 different satellite-based estimates of the imbalances of the polar ice sheets that have been derived since 1998. These studies have been of three major types - standard mass budget analyses, altimetry measurements of ice-sheet volume changes, and measurements of the ice sheets' changing gravitational attraction - and they have yielded a diversity of values, ranging from an implied sea-level rise of 1.0 mm/year to a sea-level fall of 0.15 mm/year. Based on their evaluation of these diverse findings, the two researchers come to the conclusion that the current "best estimate" of the contribution of polar ice wastage to global sea level change is a rise of 0.35 millimeters per year, which over a century amounts to only 35 millimeters, or less than an inch and a half.
Yet even this small sea level rise may be unrealistically large, for although two of Greenland's biggest outlet glaciers doubled their mass-loss rates in 2004, causing many to claim that the Greenland Ice Sheet was responding more rapidly to global warming than expected, Howat et al. (2007) report that the glaciers' mass-loss rates "decreased in 2006 to near the previous rates." And these observations, in their words, "suggest that special care must be taken in how mass-balance estimates are evaluated, particularly when extrapolating into the future, because short-term spikes could yield erroneous long-term trends."
Other findings also contradict Hansen's claim that "increasingly rapid changes on West Antarctica and Greenland ... are truly alarming." Writing in the 30 March 2007 issue of Science, for example, Anandakrishnan et al. (2007) describe a sedimentary wedge or "till delta" deposited by and under West Antarctica's Whillans Ice Stream that they detected via radar surveys made from the floating Ross Ice Shelf. This grounding-line buildup of sedimentary material, as they describe it, "serves to thicken the ice and stabilize the position of the grounding line," so that "the ice just up-glacier of the grounding line is substantially thicker than that needed to allow floatation." Consequently, they say that "the grounding-line will tend to remain in the same location ... until sea level rises enough to overcome the excess thickness that is due to the wedge."
So how high would the sea need to rise to "unground" the Whillans Ice Stream and wrest it from the continent? In a study that analyzes this question in detail, Alley et al. (2007) find that "sea-level changes of a few meters are unlikely to substantially affect ice-sheet behavior," and they conclude that a rise on the order of 100 meters might be needed to "overwhelm the stabilizing feedback from sedimentation." In fact, Anderson (2007) states that "at the current rate of sea-level rise, it would take several thousand years [our italics] to float the ice sheet off [its] bed." What is more, Alley et al. say that the ice sheet's extra thickness up-glacier from the grounding-line wedge will tend to stabilize it against "any other environmental perturbation."
With respect to the range of applicability of the findings of Anandakrishnan et al. and Alley et al., Anderson notes that "grounding-zone wedges are common features on the continental shelf, including the Ross Sea Shelf," and that "all ice streams of the Siple Coast have an anomalous elevation and stop at the grounding line," which leads him to conclude that "this mechanism for stabilization of the grounding-line is likely to be widespread." Consequently, Anderson concludes that "sea-level rise may not destabilize ice sheets as much as previously feared," which in turn suggests that sea level itself may not rise as fast or as high as previously feared. So what do actual sea level data suggest?
Sea Level TrendsLombard et al. (2005) studied temperature-induced (thermosteric) sea-level change over the last 50 years using the global ocean temperature data of Levitus et al. (2000) and Ishii et al. (2003). In doing so, they found thermosteric sea level variations are dominated by decadal oscillations of earth's major ocean-atmosphere climatic perturbations (El Niño-Southern Oscillation, Pacific Decadal Oscillation and North Atlantic Oscillation), and that thermosteric trends computed over 10-year windows exhibit 20-year oscillations with positive values of 1 to 1.5 mm/year and negative values of -1 to -1.5 mm/year. Hence, over the 50 years for which global ocean temperature data exist, there has indeed been a rise in sea level due to the thermal expansion of sea water, but only because the record begins at the bottom of a trough and ends at the top of a peak. Between these two points, there are both higher and lower values, obscuring what might be implied if earlier data were available or what may be suggested as more data are acquired. Noting further that sea level trends derived from Topex/Poseidon altimetry from 1993 to 2003 are "mainly caused by thermal expansion" and are "very likely a non-permanent feature," Lombard et al. conclude that "we simply cannot extrapolate sea level into the past or the future using satellite altimetry alone." Thus, it is to long-term coastal tide gauge records that we must turn for an evaluation of Hansen's claim that the rate of sea level rise is accelerating.
Holgate and Woodworth (2004) derived a mean global sea level history from 177 coastal tide gauge records spanning the period 1955-1998, which Holgate (2007) extended back in time another half-century by demonstrating that the combination of nine much longer high-quality tide gauge records from around the world (New York, Key West, San Diego, Balboa, Honolulu, Cascais, Newlyn, Trieste and Auckland) was similar enough to the 177-site record over the period of the two data sets' overlap to warrant the use of the nine-station record as a reasonable representation of mean global sea level for the much longer 1904-2003 period. This history is represented by the wavering black line in the figure below; and based on that history, Holgate calculated that the mean rate of global sea level rise was "larger in the early part of the last century (2.03 +/- 0.35 mm/year 1904-1953), in comparison with the latter part (1.45 +/- 0.34 mm/year 1954-2003)."
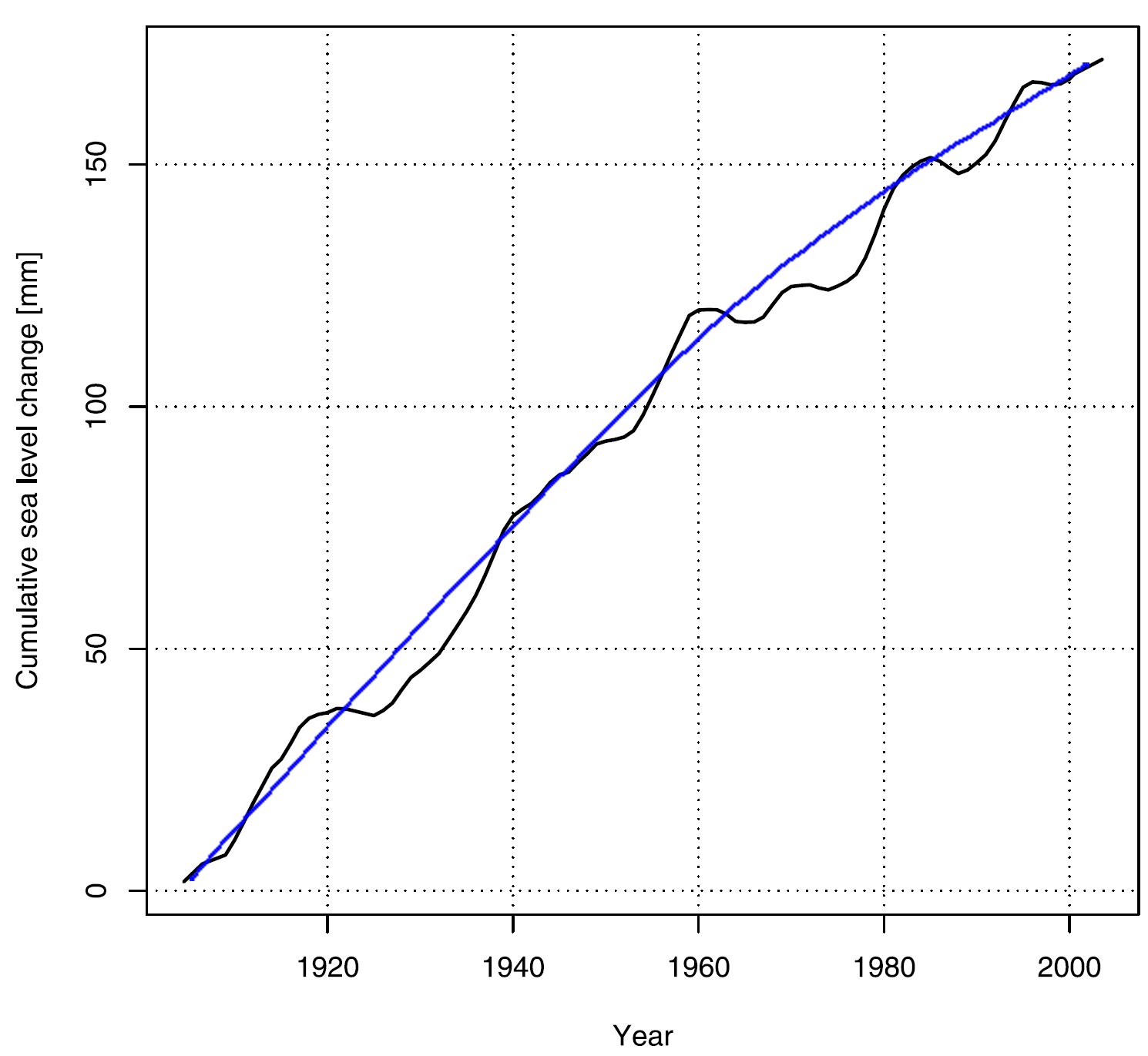
The cumulative increase in mean global sea level (1904-2003) derived from nine high-quality tide gauge records from around the world. Adapted from Holgate (2007).
Another way of thinking about Holgate's century-long sea level history is suggested by the blue curve we have fit to it, which indicates that mean global sea level may have been rising ever more slowly with the passage of time throughout the entire last hundred years. In any event, and whichever way one looks at Holgate's findings - as either two linear trends or one longer continuous curve - the nine select tide gauge records indicate that the mean rate of global sea level rise has not accelerated over the recent past. In fact, it likely has done just the opposite - in clear contradiction of Hansen's adamant claim to the contrary.
Augmenting the findings of Holgate are those of Jevrejeva et al. (2006), who analyzed information contained in the Permanent Service for Mean Sea Level database using a method based on Monte Carlo Singular Spectrum Analysis. Removing 2- to 30-year quasi-periodic oscillations, they derived nonlinear long-term trends for 12 large ocean regions, which they combined to produce the mean global sea level (gsl) and mean global sea level rate-of-rise (gsl rate) curves depicted in the figure below.
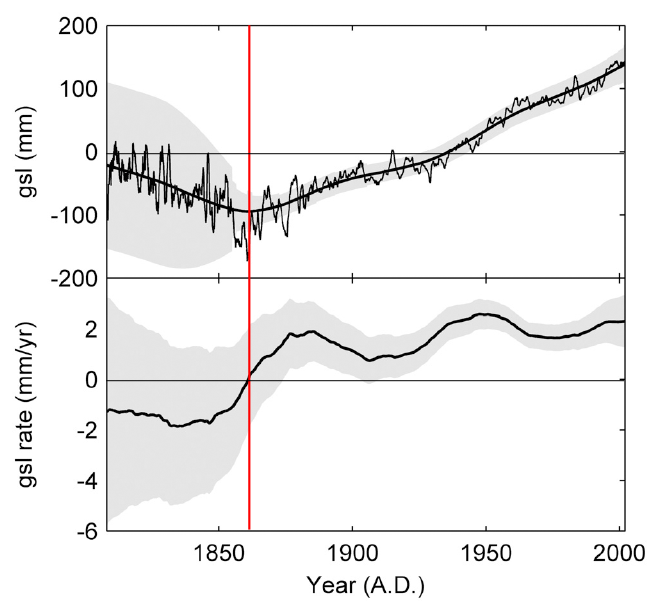
Mean gsl (top), with its shaded 95% confidence interval, and mean gsl rate (bottom), with its shaded standard error interval. Adapted from Jevrejeva et al. (2006).
In discussing their results, Jevrejeva et al. say they show that "global sea level rise is irregular and varies greatly over time," and that "it is apparent that rates in the 1920-1945 period are likely to be as large as today's." In addition, they report that their "global sea level trend estimate of 2.4 +/- 1.0 mm/year for the period from 1993 to 2000 matches the 2.6 +/- 0.7 mm/year sea level rise found from TOPEX/Poseidon altimeter data."
With respect to what Jevrejeva et al. describe as "the discussion on whether sea level rise is accelerating," their results pretty much answer the question in the negative; and in further support of this conclusion, they note that "Church et al. (2004) pointed out that with decadal variability in the computed global mean sea level, it is not possible to detect a significant increase in the rate of sea level rise over the period 1950-2000," as is clearly evident from the bottom portion of the above figure.
These observations lead us to wonder why late 20th-century global warming - which climate alarmists describe as having been unprecedented over the past two millennia or more - barely makes a ripple in the global sea level data of the two preceding figures. We are even more intrigued about the matter in light of the fact that the warming that brought an end to the Little Ice Age is readily apparent in the first, and even the second, of the three upward-trending segments of Jevrejeva et al.'s gsl rate history. Likewise, we are perplexed by the fact that the rising atmospheric CO2 concentration - which climate alarmists contend was responsible for the "unprecedented" global warming of the late 20th century - experienced a dramatic increase in its rate of rise just after 1950 (shifting from a 1900-1950 mean rate-of-rise of 0.33 ppm/year to a 1950-2000 mean rate-of-rise of 1.17 ppm/year, which is a good three and a half times greater), yet the mean global sea level rate of rise did not trend upwards after 1950, nor has it subsequently exceeded its 1950 rate-of-rise, which means that something is very wrong with the climate-alarmist theory espoused by Hansen and his dozens of collaborators.
So what does a climate modeler do when the world of nature refuses to behave as his model suggests it should? One option is to claim that the response times of many of the modeled processes are so long that the major proportions of their manifestations are yet to be seen, which enables Hansen to contend that "we have not yet felt the full climate impact of the gases that have already been added to the atmosphere," and to affirm that the predicted phenomena are still, as he says, "in the pipeline." But this argument has its problems too.
Atmospheric Methane ConcentrationsOne of the major "slow" feedback processes that Hansen identifies is "the effect of warming on emissions of long-lived greenhouse gases," such as he claims is being caused by the "melting of tundra in North America and Eurasia," which he states "is observed to be causing increased ebullition of methane from methane hydrates." The real world of nature, however, seems little impressed by these contentions; for after rising rapidly since the start of the Industrial Revolution, the air's methane concentration has been rising ever more slowly, especially during the "unprecedented" warming of the last few decades. In fact, since the beginning of the 21st century, the atmosphere's methane concentration has actually stabilized - ceasing to rise any further - as indicated by the data provided by Dlugokencky et al. (2003), which we have plotted in the figure below, and to which we have fit two linear regressions and an intervening second-order polynomial.
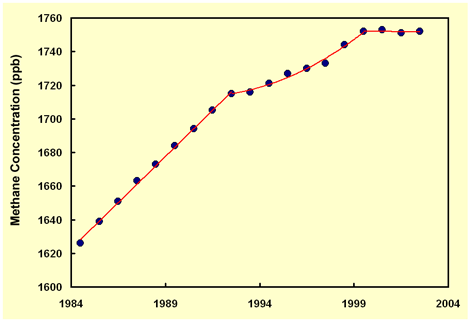
Why are these observations so important? They are important because, as Dulgokencky et al. report, "atmospheric methane's contribution to anthropogenic climate forcing is about half that from CO2 [our italics] when direct and indirect components to its forcing are summed (Hansen and Sato, 2001)." In addition, they note that "all methane emission scenarios considered by the IPCC Special Report on Emission Scenarios (Nakicenovic et al., 2000) resulted in increasing [our italics] atmospheric methane for at least the next 3 decades, and many of the scenarios projected large increases through the 21st century (Prather et al., 2001)." In reality, however, it now appears that a large portion of the anticipated global warming problem may have simply disappeared, rather than gotten much worse, as Hansen claims.
Another - and slightly expanded - perspective of the atmosphere's methane history has been presented by Khalil et al. (2007), which we have reproduced in the figure below and to which we have added the smooth green line.
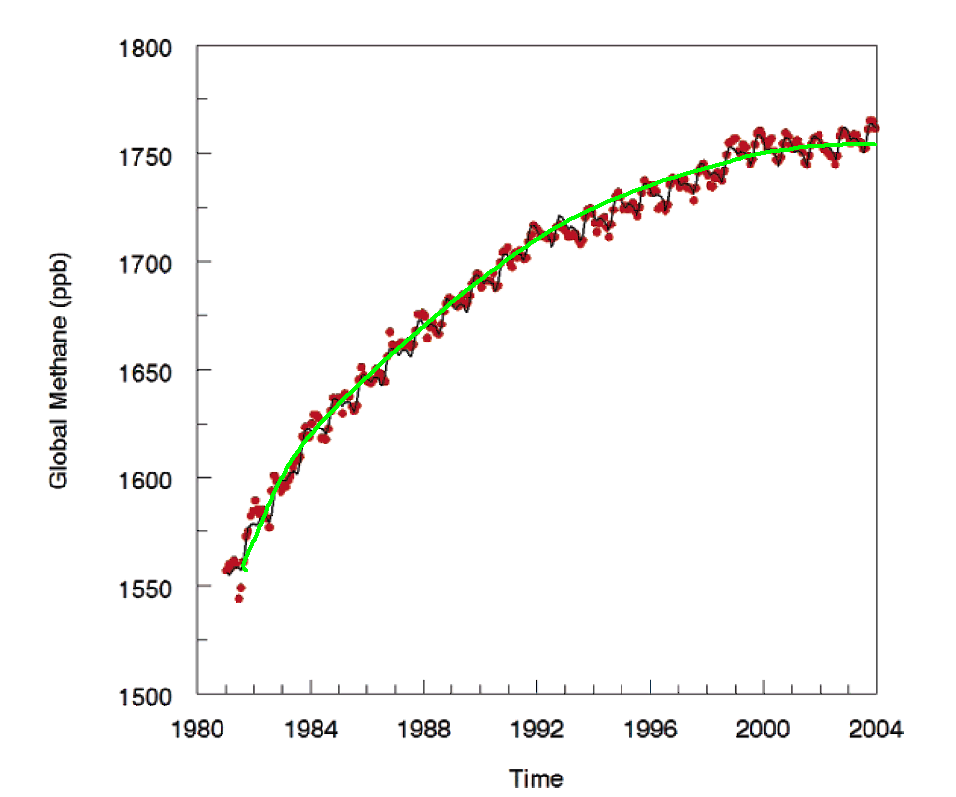
Global atmospheric methane concentration. Adapted from Khalil et al. (2007).
This graph suggests that the trend in atmospheric methane concentration, as Khalil et al. describe it, "has been decreasing for the last two decades until the present when it has reached near zero," and they say that "it is questionable whether human activities can cause methane concentrations to increase greatly in the future." In fact, there is reason to believe the global methane concentration may actually begin to decline ... and soon!
To explain the rational behind this surprising scenario, we turn to the study of Simpson et al. (2002), who presented annual global tropospheric methane growth rates for the period 1983-2000, based on measurements made by the Department of Chemistry at the University of California in Irvine, as depicted in the figure below.
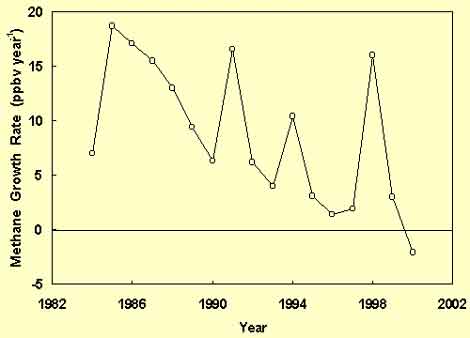
Tropospheric methane growth rate vs. time. Adapted from Simpson et al. (2002).
With respect to the data of this figure, and particularly the data from the 1990s, Simpson et al. said they "caution against viewing each year of high methane growth as an anomaly against a trend of declining methane growth." Yet that is precisely what the data suggest, i.e., a declining baseline upon which are superimposed periodic anomalous increases; and in this interpretation, we are not alone. The first of the large methane spikes depicted in the above figure is widely recognized as having been caused by the sudden eruption of Mt. Pinatubo in June of 1991 (Bekki et al., 1994; Dlugokencky et al., 1996; Lowe et al., 1997); while the last and most dramatic of the spikes has been associated with the strong El Niño of 1997-98 (Dlugokencky et al., 2001). In addition, Dlugokencky et al. (1998), Francey et al. (1999) and Lassey et al. (2000) have all felt confident in concluding the data suggest that the annual rate-of-rise of the atmosphere's methane concentration has indeed declined and led to a cessation of methane concentration growth.
Projecting ahead, if anomalous methane spikes similar to those that occurred in the 1990s continue to occur at similar intervals in the future, the atmosphere's methane concentration should continue to rise - but only very slowly - for just a few more years, after which the declining background methane growth rate, which has already turned negative, will have dropped low enough to overwhelm any short-term impacts of periodic methane spikes. At that point in time we may thus be able to see an actual decline in the air's methane concentration, which should gradually accelerate if subsequent methane spikes fail to penetrate into positive territory. Consequently, if this scenario proves to be correct, the decreasing trend in atmospheric methane concentration may soon provide a negative-greenhouse force that could counter a good deal of the positive-greenhouse force created by the ongoing rise in the air's CO2 content.
Nevertheless, and in spite of all of the real-world observations supportive of either a flat or a soon-to-be-declining trend in atmospheric methane concentration, Hansen contends in his US House of Representatives testimony that "greenhouse gases" - of which methane stands next in importance to CO2 - "are skyrocketing." In making this claim, Hansen is totally out of touch with reality.
Climates of the PastIn an attempt to depict earth's current temperature as being extremely high and, therefore, extremely dangerous, Hansen focuses almost exclusively on a single point of the earth's surface in the Western Equatorial Pacific, for which he and others (Hansen et al., 2006) compared modern sea surface temperatures (SSTs) with paleo-SSTs that were derived by Medina-Elizade and Lea (2005) from the Mg/Ca ratios of shells of the surface-dwelling planktonic foraminifer Globigerinoides rubber that they obtained from an ocean sediment core. In doing so, they concluded that "this critical ocean region, and probably the planet as a whole [our italics], is approximately as warm now as at the Holocene maximum and within ~1°C of the maximum temperature of the past million years [our italics]."
Is there any compelling reason to believe these claims of Hansen et al. about the entire planet? In a word, no, because there are a multitude of other single-point measurements that suggest something vastly different.
Even in their own paper, Hansen et al. present data from the Indian Ocean that indicate, as best we can determine from their graph, that SSTs there were about 0.75°C warmer than they are currently some 125,000 years ago during the prior interglacial. Likewise, based on data obtained from the Vostok ice core in Antarctica, another of their graphs suggests that temperatures at that location some 125,000 years ago were about 1.8°C warmer than they are now; while data from two sites in the Eastern Equatorial Pacific indicate it was approximately 2.3 to 4.0°C warmer compared to the present at about that time. In fact, Petit et al.'s (1999) study of the Vostok ice core demonstrates that large periods of all four of the interglacials that preceded the Holocene were more than 2°C warmer than the peak warmth of the current interglacial.
But we don't have to go nearly so far back in time to demonstrate the non-uniqueness of current temperatures. Of the five SST records that Hansen et al. display, three of them indicate the mid-Holocene was also warmer than it is today. Indeed, it has been known for many years that the central portion of the current interglacial was much warmer than its latter stages have been. To cite just a few examples of pertinent work conducted in the 1970s and 80s - based on temperature reconstructions derived from studies of latitudinal displacements of terrestrial vegetation (Bernabo and Webb, 1977; Wijmstra, 1978; Davis et al., 1980; Ritchie et al., 1983; Overpeck, 1985) and vertical displacements of alpine plants (Kearney and Luckman, 1983) and mountain glaciers (Hope et al., 1976; Porter and Orombelli, 1985) - we note it was concluded by Webb et al. (1987) and the many COHMAP Members (1988) that mean annual temperatures in the Midwestern United States were about 2°C greater than those of the past few decades (Bartlein et al., 1984; Webb, 1985), that summer temperatures in Europe were 2°C warmer (Huntley and Prentice, 1988) - as they also were in New Guinea (Hope et al., 1976) - and that temperatures in the Alps were as much as 4°C warmer (Porter and Orombelli, 1985; Huntley and Prentice, 1988). Likewise, temperatures in the Russian Far East are reported to have been from 2°C (Velitchko and Klimanov, 1990) to as much as 4-6°C (Korotky et al., 1988) higher than they were in the 1970s and 80s; while the mean annual temperature of the Kuroshio Current between 22 and 35°N was 6°C warmer (Taira, 1975). Also, the southern boundary of the Pacific boreal region was positioned some 700 to 800 km north of its present location (Lutaenko, 1993).
But we needn't go back to even the mid-Holocene to encounter warmer-than-present temperatures, as the Medieval Warm Period, centered on about AD 1100, had lots of them. In fact, every single week since 1 Feb 2006, we have featured on our website (www.co2science.org) a different peer-reviewed scientific journal article that testifies to the existence of this several-centuries-long period of notable warmth, in a feature we call our Medieval Warm Period Record of the Week. Also, whenever it has been possible to make either a quantitative or qualitative comparison between the peak temperature of the Medieval Warm Period (MWP) and the peak temperature of the Current Warm Period (CWP), we have included those results in the appropriate quantitative or qualitative frequency distributions we have posted within this feature; and a quick perusal of these ever-growing databases (reproduced below as of 23 May 2007) indicates that, in the overwhelming majority of cases, the peak warmth of the Medieval Warm Period was significantly greater than the peak warmth of the Current Warm Period.
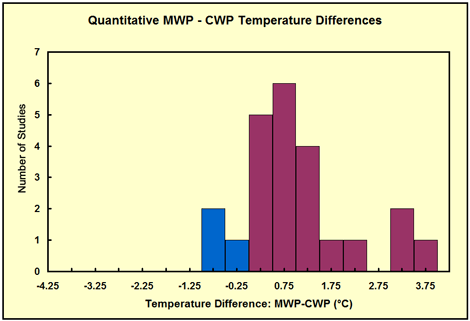
The distribution in 0.5°C increments of Level 1 Studies that allow one to identify the degree by which peak MWP temperatures either exceeded (positive values, red) or fell short of (negative values, blue) peak CWP temperatures.
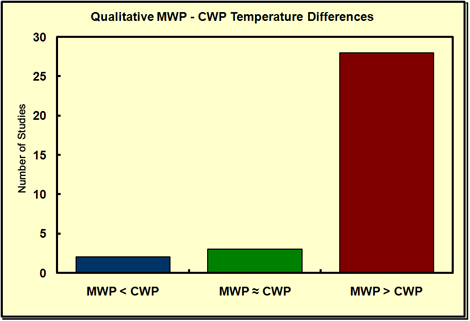
The distribution of Level 2 Studies - not including Level 1 Studies - that allow one to determine whether peak MWP temperatures were warmer than (red), equivalent to (green), or cooler than (blue), peak CWP temperatures.
In concluding this portion of our critique of Hansen's testimony, we note that the mean surface air temperature of the earth is currently nowhere near as high as it was a million years ago. Neither are current temperatures as high as the peak temperatures of the prior four interglacials, nor are they as high as they were during the central portion of the current interglacial. In fact, it's not even as warm now as it was a paltry 900 years ago, when the atmosphere's CO2 concentration was 100 ppm less than it is today, which sure doesn't say much for the warming power of CO2 nor for the storyline promoted in Hansen's testimony.
Warming-Induced Extinctions of Terrestrial Plants and AnimalsHansen writes that "continued business-as-usual greenhouse gas emissions threaten many ecosystems," contending - even more ominously - that "very little additional [climate] forcing is needed ... to cause the extermination of a large fraction of plant and animal species." But where is the evidence for these claims? Hansen says that "animals and plants migrate as climate changes," and so they do, both upward in altitude and poleward in latitude; and he states that in response to global warming, "polar species can be pushed off the planet [i.e., driven to extinction], as they have no place else to go," and that "life in alpine regions ... is similarly in danger of being pushed off the planet." But again, where is the evidence to support these contentions?
In searching Hansen's testimony and his "accepted for publication" manuscript on the subject, we could find no real-world support for this aspect of his climate-alarmist thesis. What we did find was typically of the same nature as Hansen's own writings: claims, contentions and opinions, but no hard evidence. Such is also the case with many peer-reviewed science journal articles that promote the same philosophy, such as those of Root et al. (2003) and Parmesan and Yohe (2003). However, as we have indicated in a major study of the topic that is archived on our website (Idso et al., 2003), even these studies have failed to provide any hard data in support of their egregious extrapolations.
So what's the real situation with respect to rising air temperatures and atmospheric CO2 concentrations, as well as the life-and-death impacts they may - or may not - have on earth's plants and animals?
A good place to begin in answering this question is with the growth-enhancing effects of elevated atmospheric CO2, which typically increase with rising air and leaf temperatures. This phenomenon is illustrated by the data of Jurik et al. (1984), who exposed bigtooth aspen leaves to atmospheric CO2 concentrations of 325 and 1935 ppm and measured their photosynthetic rates at a number of different temperatures. In the figure below, we have reproduced their results and slightly extended the two relationships defined by their data to both warmer and cooler conditions.
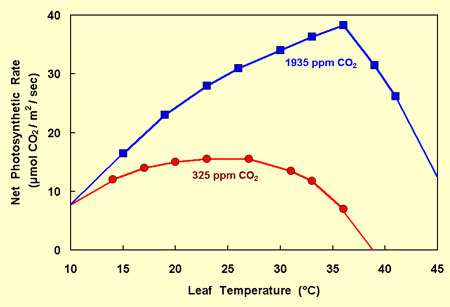
In viewing this figure, it can be seen that at a leaf temperature of 10°C, elevated CO2 has essentially no effect on net photosynthesis in this particular species, as Idso and Idso (1994) have demonstrated is characteristic of plants in general. At 25°C, however, where the net photosynthetic rate of the leaves exposed to 325 ppm CO2 is maximal, the extra CO2 of this study boosted the net photosynthetic rate of the foliage by nearly 100%; and at 36°C, where the net photosynthetic rate of the leaves exposed to 1935 ppm CO2 is maximal, the extra CO2 boosted the net photosynthetic rate of the foliage by a whopping 450%. In addition, the extra CO2 increased the optimum temperature for net photosynthesis in this species by about 11°C: from 25°C in air of 325 ppm CO2 to 36°C in air of 1935 ppm CO2.
In viewing the warm-temperature projections of the two relationships at the right-hand side of the figure, it can additionally be seen that the transition from positive to negative net photosynthesis - which denotes a change from life-sustaining to life-sapping conditions - likely occurs somewhere in the vicinity of 39°C in air of 325 ppm CO2 but somewhere in the vicinity of 50°C in air of 1935 ppm CO2. Consequently, not only was the optimum temperature for photosynthesis of bigtooth aspen greatly increased by the extra CO2 of this experiment, so too was the lethal temperature (above which life cannot long be sustained) likewise increased, and by approximately the same amount, i.e., 11°C.
These observations, which are similar to what has been observed in many other plants, suggest that when the atmosphere's temperature and CO2 concentration rise together (Cowling, 1999), the vast majority of earth's plants would likely not feel a need (or only very little need) to migrate towards cooler regions of the globe. Any warming would obviously provide them an opportunity to move into places that were previously too cold for them, but it would not force them to move, even at the hottest extremes of their ranges; for as the planet warmed, the rising atmospheric CO2 concentration would work its biological wonders, significantly increasing the temperatures at which most of earth's C3 plants - which comprise about 95% of the planet's vegetation - function best, creating a situation where earth's plant life would actually "prefer" warmer conditions.
So what do we find at the tops of alpine mountains nowadays? Have any plants there been "pushed off the planet" in response to supposedly unprecedented 20th-century global warming?
Walther et al. (2005) investigated this climate-alarmist nightmare by resurveying (in July/August 2003) the floristic composition of the uppermost ten meters of ten mountain summits in the Swiss Alps, applying the same methodology used in earlier surveys of the same mountain tops by Rubel (1912), which was conducted in 1905, and Hofer (1992), which was conducted in 1985. Hence, their analysis covered the bulk of the Little Ice Age-to-Current Warm Period transition (1905-2003), the last portion of which (1985-2003) is claimed by climate alarmists such as Hansen to have experienced a warming that was unprecedented over the past two millennia (or more!) in terms of both the rate of temperature rise and the degree to which the temperature rose.
This work revealed that plants of many species marched up the mountainsides of the Swiss Alps as the earth warmed, but that none of them were "pushed off the planet." As a result, the species richness (i.e., biodiversity) of the ten mountaintops was dramatically increased over the past century of global warming. For the time interval 1905-1985, for example, the mean increase in species numbers recorded by Hofer (1992) was 86%; and Walther et al. report that "species numbers recorded in 2003 were generally more than double (138%) compared to the results by Rubel (1912) and 26% higher than those reported by Hofer (1992)." Put another way, they say that "the rate of change in species richness (3.7 species/decade) was significantly greater in the later period compared to the Hofer resurvey (1.3 species/decade)." Most important of all, they say that "the observed increase in species numbers does not entail the replacement of high alpine specialists by species from lower altitudes [our italics], but rather has led to an enrichment [our italics] of the overall summit plant diversity."
Another pertinent study of evolving mountaintop biodiversity was conducted by Kullman (2007), who analyzed the changing behavior of alpine and subalpine plants, together with shifts in their geographical patterns, during the past century, when air temperatures rose by about 1°C in the Scandes of west-central Sweden, which "methodical approach," in his words, "also included repeat photography, individual age determinations and analyses of permanent plots." This work revealed, according to Kullman, that "at all levels, from trees to tiny herbs, and from high to low altitudes, the results converge to indicate a causal association between temperature rise and biotic evolution." More specifically, he reports that "treeline advance since the early 20th century varies between 75 and 130 m, depending on species and site," and that "subalpine/alpine plant species have shifted upslope by [an] average [of] 200 m." In addition, he states that "present-day repetitions of floristic inventories on two alpine mountain summits reveal increases of plant species richness by 58 and 67%, respectively, since the early 1950s." And again, Kullman also reports that "no species have yet become extinct from the highest elevations [our italics]," adding that his results "converge with observations in other high-mountain regions worldwide," in support of which statement he cites the studies of Grabherr et al. (1994), Keller et al. (2000), Kullman (2002), Virtanen et al. (2003), Klanderud and Birks (2003), Walther et al. (2005) and Lacoul and Freedman (2006).
Switching from plants to animals, Parmesan et al. (1999) examined the distributional changes, broadly spread over the past century, of non-migratory species of butterflies whose northern boundaries were in northern Europe and whose southern boundaries were in southern Europe or northern Africa. Their analysis indicated that the northern boundaries of the ranges of 52 species shifted northward for 65% of them, remained stable for 34% of them, and shifted southward for 2% of them, while the southern boundaries of the ranges of 40 species shifted northward for 22% of them, remained stable for 72% of them, and shifted southward for 5% of them. Consequently, in the words of the thirteen researchers who conducted the work, "nearly all northward shifts involved extensions at the northern boundary with the southern boundary remaining stable."
Since this is precisely the type of behavior we would expect for plants in a CO2-enriched and warming world - i.e., an opportunity for significant poleward expansion at the cold edge of a species' range, but little to no impetus for poleward migration at the warm edge of its range - it is possible that the observed changes in European butterfly ranges over the past century of concomitant warming and rising atmospheric CO2 concentration are related to matching changes in the ranges of the plants upon which the butterflies depend for food. Or, the similarity could be due to some more complex phenomenon, possibly even a direct physiological effect of temperature and atmospheric CO2 concentration at work on the butterflies themselves.
In any event, in the face of the 0.8°C of warming experienced in Europe over the 20th century and the 75-ppm (25%) increase in atmospheric CO2 concentration experienced concurrently, the ultimate consequence for European butterflies has not been threatening at all, much less a portent of extinction. In fact, since "nearly all northward [range] shifts involved extensions at the northern boundary with the southern boundary remaining stable," according to Parmesan et al., "most species effectively expanded the size of their range when shifting northwards," which likely strengthened them against the possibility of extinction.
Although we have highlighted the findings of just a few real-world studies of the effects of concomitant increases in air temperature and atmospheric CO2 concentration on the "sustainability" of earth's plant and animal species, many additional studies that have yielded similar results have been described in detail by Idso et al. (2003), whose report on the subject can be found on our website and should be considered a vital appendage of our critique of Hansen's testimony. Consequently - and not wanting to "beat a dead horse" any further in this regard - we proceed to a consideration of a woefully under-reported aspect of the topic that is almost never discussed in the climate-alarmist literature that portrays anthropogenic CO2 emissions as leading to massive species extinctions. And why does it fail to appear there, as well as in Hansen's testimony? It fails to appear because this aspect of the subject totally undercuts climate-alarmist policy prescriptions for averting their contrived species extinction catastrophe, as well as the many other climate-related disasters described by Hansen.
The CO2-Induced Preservation of Terrestrial SpeciesHow much land can ten billion people spare for nature? This provocative question was posed by Waggoner (1995) in an insightful essay wherein he explored the dynamic tension that exists between the need for land to support the agricultural enterprises that sustain mankind, and the need for land to support the natural ecosystems that sustain all other creatures. This challenge of meeting our future food needs - and not decimating the rest of the biosphere in the process - was stressed even more strongly by Huang et al. (2002), who wrote that humans "have encroached on almost all of the world's frontiers, leaving little new land that is cultivatable." And in consequence of humanity's usurpation of this most basic of natural resources, Raven (2002) stated in his Presidential Address to the American Association for the Advancement of Science that "species-area relationships, taken worldwide in relation to habitat destruction, lead to projections of the loss of fully two-thirds of all species on earth by the end of this century."
In a more detailed analysis of the nature and implications of this impending "global land-grab" - which moved it closer to the present by a full half-century - Tilman et al. (2001) concluded that the task of meeting the doubled world food demand, which they calculated would exist in the year 2050, would likely exact a toll that "may rival climate change in environmental and societal impacts." But how could something so catastrophic manifest itself so soon?
Tilman and his nine collaborators shed some light on this question by noting that at the end of the 20th century mankind was already appropriating "more than a third of the production of terrestrial ecosystems and about half of usable freshwaters." Now, think of doubling those figures, in order to meet the doubled global food demand that Tilman et al. predict for the year 2050. The results suggest that a mere 43 years from now mankind will be appropriating more than two thirds of terrestrial ecosystem production plus all of earth's remaining usable freshwater, as has also been discussed by Wallace (2000).
In terms of land devoted to agriculture, Tilman et al. calculate a much less ominous 18% increase by the year 2050. However, because most developed countries are projected to withdraw large areas of land from farming over the next fifty years, the loss of natural ecosystems to crops and pastures in developing countries will amount to about half of their remaining suitable land, which would, in the words of the Tilman team, "represent the worldwide loss of natural ecosystems larger than the United States." What is more, they say that these land usurpations "could lead to the loss of about a third of remaining tropical and temperate forests, savannas, and grasslands." And in a worrisome reflection upon the consequences of these land-use changes, they remind us that "species extinction is an irreversible impact of habitat destruction."
What can be done to avoid this horrific situation? In a subsequent analysis, Tilman et al. (2002) introduced a few more facts before suggesting some solutions. First of all, they noted that by 2050 the human population of the globe is projected to be 50% larger than it was just prior to the writing of their paper, and that global grain demand by 2050 could well double, due to expected increases in per capita real income and dietary shifts toward a higher proportion of meat. Hence, they but stated the obvious when they concluded that "raising yields on existing farmland is essential for 'saving land for nature'."
So how can this readily-defined but Herculean task be accomplished? Tilman et al. proposed a strategy that focuses on three essential efforts: (1) increasing crop yield per unit of land area, (2) increasing crop yield per unit of nutrients applied, and (3) increasing crop yield per unit of water used.
With respect to the first of these efforts - increasing crop yield per unit of land area - the researchers note that in many parts of the world the historical rate-of-increase in crop yield is declining, as the genetic ceiling for maximal yield potential is being approached. This observation, in their estimation, "highlights the need for efforts to steadily increase the yield potential ceiling." With respect to the second effort - increasing crop yield per unit of nutrients applied - they note that "without the use of synthetic fertilizers, world food production could not have increased at the rate [that it did in the past] and more natural ecosystems would have been converted to agriculture." Hence, they say that the ultimate solution "will require significant increases in nutrient use efficiency, that is, in cereal production per unit of added nitrogen." Finally, with respect to the third effort - increasing crop yield per unit of water used - Tilman et al. note that "water is regionally scarce," and that "many countries in a band from China through India and Pakistan, and the Middle East to North Africa either currently or will soon fail to have adequate water to maintain per capita food production from irrigated land." Increasing crop water use efficiency, therefore, is also a must.
Although the impending man vs. nature crisis and several important elements of its potential solution are thus well defined, Tilman and his first set of collaborators concluded that "even the best available technologies, fully deployed, cannot prevent many of the forecasted problems." This was also the finding of Idso and Idso (2000), who concluded that although "expected advances in agricultural technology and expertise will significantly increase the food production potential of many countries and regions," these advances "will not increase production fast enough to meet the demands of the even faster-growing human population of the planet."
How can we prevent this unthinkable catastrophe from occurring, especially when it has been concluded by highly-credentialed researchers that earth possesses insufficient land and freshwater resources to forestall it, while simultaneously retaining any semblance of the natural world and its myriad animate creations? Although the task may appear next to impossible to accomplish, it can be done; for we have a powerful ally in the ongoing rise in the atmosphere's CO2 concentration that can provide what we can't.
Since atmospheric CO2 is the basic "food" of nearly all plants, the more of it there is in the air, the better they function and the more productive they become. For a 300-ppm increase in the atmosphere's CO2 concentration above the planet's current base level of slightly less than 400 ppm, for example, the productivity of earth's herbaceous plants rises by something on the order of 30% (Kimball, 1983; Idso and Idso, 1994), while the productivity of its woody plants rises by something on the order of 50% (Saxe et al., 1998; Idso and Kimball, 2001). Thus, as the air's CO2 content continues to rise, so too will the productive capacity or land-use efficiency of the planet continue to rise, as the aerial fertilization effect of the upward-trending atmospheric CO2 concentration boosts the growth rates and biomass production of nearly all plants in nearly all places. In addition, elevated atmospheric CO2 concentrations typically increase plant nutrient-use efficiency in general - and nitrogen-use efficiency in particular - as well as plant water-use efficiency, as may be verified by perusing the many reviews of scientific journal articles we have produced on these topics and archived in the Subject Index of our website (www.co2science.org). Consequently, with respect to fostering all three of the plant physiological phenomena that Tilman et al. (2002) contend are needed to prevent the catastrophic consequences they foresee for the planet just a few short decades from now, a continuation of the current upward trend in the atmosphere's CO2 concentration would appear to be essential.
In the case we are considering here, for example, the degree of crop yield enhancement likely to be provided by the increase in atmospheric CO2 concentration expected to occur between 2000 and 2050 has been calculated by Idso and Idso (2000) to be sufficient - but only by the slightest of margins - to compensate for the huge differential that is expected to otherwise prevail between the supply and demand for food earmarked for human consumption just 43 years from now. Consequently, letting the evolution of technology take its natural course, with respect to anthropogenic CO2 emissions, would appear to be the only way we will ever be able to produce sufficient agricultural commodities to support ourselves in the year 2050 without the taking of unconscionable amounts of land and freshwater resources from nature and decimating the biosphere in the process.
But what about life in the oceans?
CO2-Induced Extinctions of Calcifying Marine OrganismsFor some time now the ongoing rise in the atmosphere's CO2 concentration has been predicted by climate alarmists to raise havoc with earth's coral reefs and other calcifying marine organisms by acidifying the world's oceans and thus lowering the calcium carbonate saturation state of seawater, making it ever more difficult for these creatures to produce their calcium carbonate skeletons; and in this regard, Hansen claims "we will be able to avoid acidification of the ocean with its destruction of coral reefs and other ocean life" if we follow his policy prescriptions. However, there is no compelling reason to believe that "coral reefs and other ocean life" will be significantly harmed - much less "destroyed" - by continuing to let technology take its natural course in terms of transitioning from fossil fuel-burning to other forms of energy production; for just like the CO2-induced global warming concept itself, the CO2-induced acidification of the world's oceans - and especially its deadly consequences concept - is an unproven theoretical construct that ignores many important biological phenomena. Nevertheless, the degree to which this catastrophic concept of CO2-induced death-in-the-oceans has been embraced, even by scientists, is nothing short of astounding, as is indicated by a paper authored by 27 researchers from eight countries that was published in the 29 September 2005 issue of Nature (Orr et al., 2005), in which the group wrote that under a "business-as-usual" scenario of future anthropogenic CO2 emissions, "key marine organisms - such as corals and some plankton - will have difficulty maintaining their external calcium carbonate skeletons," and where they suggested that these dire conditions "could develop within decades, not centuries as suggested previously."
So what's the story here? Is there any real-world evidence that can be cited in support of these strident claims? Hansen and Orr et al. certainly make it appear that such exists, but a little scientific sleuthing reveals nothing of substance that supports their claim. In fact, it actually suggests just the opposite of what they predict.
We begin by noting the 27 scientists contend that (1) in response to the ongoing rise in the air's CO2 content, "aqueous CO2 concentrations will increase and carbonate ion concentrations will decrease, making it more difficult for marine calcifying organisms to form biogenic calcium carbonate," and that (2) "substantial experimental evidence indicates that calcification rates will decrease in low-latitude corals (Millero, 1995; Dickson, 1990; Dickson and Riley, 1979), which form reefs out of aragonite [a metastable form of calcium carbonate (CaCO3)], and in phytoplankton that form their tests (shells) out of calcite (Mucci, 1983; Bischoff et al., 1987), the stable form of CaCO3)." In reviewing the five papers they cite in support of these contentions, however, we find that none of them deal with living organisms, and, therefore, that none of them deal with the calcification process as it is conducted in nature by living entities.
We have previously written extensively about the importance of not excluding life from such important considerations, noting that calcification is much more than a physical-chemical process that can be accurately described by a set of equations. Rather, we have emphasized, time and again, that coral calcification is a biologically-driven physical-chemical process that may not yet be amenable to explicit mathematical description. In fact, we reported several years ago (Idso et al., 2000) - based on proper citations of the scientific literature - that "the photosynthetic activity of zooxanthellae is the chief source of energy for the energetically-expensive process of calcification," and that considerable evidence shows that "long-term reef calcification rates generally rise in direct proportion to increases in rates of reef primary production," which suggests that if anthropogenic-induced increases in the transfer of CO2 from the air to the world's oceans were to lead to increases in coral symbiont photosynthesis - as atmospheric CO2 enrichment generally does for nearly all land plants - it is likely that increases in coral calcification rates would occur as well.
We have also noted that the calcium carbonate saturation state of seawater actually rises with an increase in temperature, countering the adverse oceanic chemistry consequences of an increase in aqueous CO2 concentration, which is a matter that is also considered by Orr et al., but which they dismiss as having a rather small effect, "typically counteracting less than 10% of the decrease due to the geochemical effect." With this little problem thus handily dispatched - and ignoring the many ways in which the forces of life might enter the picture - they calculate that "relative to preindustrial conditions, invasion of anthropogenic CO2 has already reduced modern surface carbonate ion concentrations by more than 10%," and they further calculate - "in agreement with previous predictions (Kleypas et al., 1999)" - that a 45% reduction relative to preindustrial levels may be reached by the end of the century, and that ultimately, "rates of calcification could decline even further, to zero." We, however, suggest these contentions are grossly in error.
So what do real-world studies of living and fossil corals and phytoplankton reveal about the various claims and counterclaims swirling about this issue? Have the increases in air temperature and atmospheric CO2 concentration that have occurred since the beginning of the Industrial Revolution seriously hampered coral and phytoplankton calcification rates? Let's review what some actual observations have to say about the matter.
In a study of calcification rates of massive Porites coral colonies on Australia's Great Barrier Reef (GBR), Lough and Barnes (1997) found that "the 20th century has witnessed the second highest period [our italics] of above average calcification in the past 237 years." Intrigued by this observation, the two researchers subsequently analyzed the calcification characteristics of 245 similar-sized massive colonies of Porites corals obtained from 29 sites located along the length, and across the breadth, of the GBR, which data spanned a latitudinal range of approximately 9° and an annual average sea surface temperature (SST) range of 25-27°C (Lough and Barnes, 2000). To these data they then added other published data from the Hawaiian Archipelago (Grigg, 1981, 1997) and Phuket, Thailand (Scoffin et al., 1992), thereby extending the latitudinal range of their expanded data set to 20° and the annual average SST range to 23-29°C.
This analysis indicated that GBR calcification rates were linearly related to average annual SST, such that "a 1°C rise in average annual SST increased average annual calcification by 0.39 g cm-2 year-1." Results were much the same for the extended data set they developed; and they report that "the regression equation explained 83.6% of the variance in average annual calcification," while noting that "this equation provides for a change [increase] in calcification rate of 0.33 g cm-2 year-1 for each 1°C change [increase] in average annual SST," in spite of unprecedented concurrent increases in atmospheric CO2 concentration.
Noting that their results "allow assessment of possible impacts of global climate change on coral reef ecosystems," Lough and Barnes determined that between the two 50-year periods 1880-1929 and 1930-1979, there was a mean calcification increase of 0.06 g cm-2 year-1; and they note that "this increase [our italics] of ~4% in calcification rate conflicts with the estimated decrease [our italics] in coral calcification rate of 6-14% over the same time period suggested by Kleypas et al. (1999) as a response to changes in ocean chemistry." Even more stunning was their observation that between the two 20-year periods 1903-1922 and 1979-1998, the warming-induced increase in calcification was about 12% in the central GBR, about 20% in the southern GBR and as much as 50% to the south of the GBR. In light of these real-world observations, therefore, and in stark contrast to the predictions of Kleypas et al. (1999), Orr et al. (2005) and the testimony of Hansen, the two researchers concluded that coral calcification rates "may have already significantly increased [our italics] along the GBR in response to global climate change."
Two other scientists that investigated the subject by means of real-world data were Bessat and Buigues (2001), who worked with a core retrieved from a massive Porites coral on the French Polynesian island of Moorea that covered the period 1801-1990, and who said they undertook the study because they thought it "may provide information about long-term variability in the performance of coral reefs, allowing unnatural changes to be distinguished from natural variability." This effort revealed that a 1°C increase in water temperature increased coral calcification rate by 4.5%, and that "instead of a 6-14% decline in calcification over the past 100 years computed by the Kleypas group, the calcification has increased." And to further emphasize this point, they reiterated that their results "do not confirm those predicted by the Kleypas et al. (1999) model," which is merely an earlier version of the Orr et al. model.
Nevertheless, and in spite of these real-world observations that refute the "lifeless" worldview of Kleypas et al. and Orr et al., certain researchers such as Buddemeier et al. (2004) have continued to claim that the ongoing rise in the air's CO2 content, and its predicted ability to lower surface ocean water pH, could dramatically decrease coral calcification rates, which they claim could lead to "a slow-down or reversal of reef-building and the potential loss of reef structures in the future." However, they were forced to admit - and in the very same publication - that "temperature and calcification rates are correlated," and that the corals of the real-world "have so far responded more to increases in water temperature (growing faster through increased metabolism and the increased photosynthetic rates of their zooxanthellae) than to decreases in carbonate ion concentration."
At about the same time, and following in the footsteps of Lough and Barnes who had worked in the Indo-Pacific, Carricart-Ganivet (2004) developed relationships between coral calcification rates and annual average SSTs based on data collected from colonies of the reef-building coral Montastraea annularis at twelve locations in the Gulf of Mexico and the Caribbean Sea. This work revealed, in his words, that "calcification rate in the Gulf of Mexico increased 0.55 g cm-2 year-1 for each 1°C increase, while, in the Caribbean Sea, it increased 0.58 g cm-2 year-1 for each 1°C increase," a result that was nearly twice as great as that obtained by Lough and Barnes for Porites corals. Further pooling these data "with those of M. annularis and M. faveolata, growing up to 10 m depth in Carrie Bow Cay, Belize, reported by Graus and Macintyre (1982), those of Dodge and Brass (1982) from all the reefs they studied at St. Croix, US Virgin Islands, and those of M. faveolata, growing up to 10 m depth in Curacao, Netherlands, Antilles, reported by Bosscher (1993)," Carricart-Ganivet obtained a relationship of ~0.5 g cm-2 year-1 for each 1°C increase in annual average SST.
To these papers can be added many others that also depict increasing coral calcification rates in the face of rising temperatures and/or atmospheric CO2 concentrations, including those of Clausen and Roth (1975), Coles and Coles (1977), Kajiwara et al. (1995), Nie et al. (1997), Reynaud-Vaganay et al. (1999) and Reyanud et al. (2004). As for why this is the way earth's corals respond, McNeil et al. (2004) say that "observed increases in coral reef calcification with ocean warming are most likely due to an enhancement in coral metabolism and/or increases in photosynthetic rates of their symbiotic algae," as we have consistently said when noting over and over that coral calcification is a biologically-driven process that can overcome physical-chemical limitations that in the absence of life would appear to be insurmountable.
Another reason for not believing that the ongoing rise in the atmosphere's CO2 content will lead to reduced oceanic pH and, therefore, lower calcification rates in the world's coral reefs and other calcifying organisms, is that the same phenomenon that powers the twin processes of coral calcification and phytoplanktonic growth (i.e., photosynthesis) tends to increase the pH of marine waters (Gnaiger et al., 1978; Santhanam et al., 1994; Brussaard et al., 1996; Lindholm and Nummelin, 1999; Macedo et al., 2001; Hansen, 2002); and this phenomenon has been shown to have the ability to dramatically increase the pH of marine bays, lagoons and tidal pools (Gnaiger et al., 1978; Santhanam, 1994; Macedo et al., 2001; Hansen, 2002), as well as significantly enhance the surface water pH of areas as large as the North Sea (Brussaard et al., 1996).
Before concluding our discussion of this important subject, however, we briefly switch our focus from corals to phytoplankton, beginning with a review of the work of Riebesell (2004), who notes that "doubling present-day atmospheric CO2 concentrations is predicted to cause a 20-40% reduction in biogenic calcification of the predominant calcifying organisms, the corals, coccolithophorids, and foraminifera." In a significant challenge to this climate-alarmist dogma, Riebesell notes that a moderate increase in CO2 actually facilitates photosynthetic carbon fixation of certain phytoplankton, such as the coccolithophorids, as represented by Emiliania huxleyi and Gephyrocapsa oceanica. In fact, Riebesell writes that "CO2-sensitive taxa, such as the calcifying coccolithophorids, should therefore benefit more [our italics] from the present increase in atmospheric CO2 compared to the non-calcifying diatoms."
More recently, Crabbe et al. (2006) used digital photography, image analysis and measurements in the field to determine the original growth rates of long-dead Quaternary corals found in exposed onshore limestone deposits near the margins of Hoga and Kaledupa Islands in the Wakatobi Marine National Park of Indonesia, after which they compared them to the growth rates of present-day corals of the same genera (Porites and Favites) living in the same area. This work revealed that the Quaternary corals appeared to have grown "in a comparable environment to modern reefs at Kaledupa and Hoga," except, of course, for the air's CO2 concentration, which is currently higher than it has been at any other time throughout the entire Quaternary, i.e., the past 1.8 million years. In addition, their measurements indicated that the radial growth rates of the modern corals were 31% greater than those of their more ancient Quaternary cousins in the case of Porites species, and 34% greater in the case of Favites species. Clearly, therefore, the impact of the historical increase in the atmosphere's CO2 concentration on the corals in question has not been as catastrophically negative as Hansen suggests it should have been. In fact, the increase in the CO2 content of the modern atmosphere appears to have not been negative at all. In fact, it appears to have been positive.
Most interesting of all, perhaps, Fine and Tchernov (2007) grew 30 coral fragments from five colonies of the scleractinian Mediterranean species Oculina patagonica and Madracis pharencis within indoor flow-through systems under ambient Mediterranean seawater temperatures and photoperiod in water maintained at pH values of 7.3-7.6 (acidified) and 8.0-8.3 (ambient) for a period of 12 months. After one month in the acidic conditions, they report there was an elongation of the coral polyps that was "followed by dissociation of the colony form and complete skeleton dissolution." However, they observed that "the polyps remained attached to the undissolved hard rocky substrate." In fact, they found that "the biomass of the solitary polyps under acidic conditions was three times as high [our italics] as the biomass of the polyps in the control colonies that continued to calcify." In addition, they say that both "control and treatment fragments maintained their algal symbionts during the entire experiment, except for six fragments (10%) of O. patagonica that partially lost their symbionts (bleached) during July but recovered within 2 months." And they report that "after 12 months, when transferred back to ambient pH conditions, the experimental soft-bodied polyps calcified and reformed colonies [our italics]." Thus, after restating their major finding that "in the absence of conditions supporting skeleton building, both species maintained basic life functions as skeleton-less ecophenotypes," Fine and Tchernov concluded that "corals might survive large-scale environmental change, such as that expected for the following century." And why not? If they've done it before - as some have theorized (Stanley and Fautin, 2001; Stanley, 2003; Medina et al., 2006), and as Fine and Tchernov have actually demonstrated can in truth be done - they likely have the capacity do it again ... and again ... and again.
Miscellaneous Misstatements of HansenIn addition to the major misconceptions he promulgates with respect to the several subjects we discus above, Hansen makes a number of brief but serious misstatements throughout his testimony, the most serious of which we identify and discuss in what follows.
(1) Hansen says "earth's history shows that climate is remarkably sensitive to global forcings" and that "positive feedbacks predominate," causing "the entire planet to be whipsawed between climatic states." Exercising but a modicum of thought, however, it is readily realized that the three parts of this unqualified contention are not universally-accepted facts, as Hansen makes them out to be, but merely opinions, and arguable ones at that. Just as easily, for example, one could say that earth's climate is not strongly influenced by global forcings and that negative feedbacks predominate, allowing the bulk of the planet to never stray too far from a climatic state conducive to the continued existence of life. And as compelling evidence for the veracity of this latter view, one need only observe the mind-boggling diversity and total number of plant and animal species that currently inhabit the planet, plus the fact that they have all been around for a very long time. How could this vast assemblage of life possibly exist today, if earth's climate was truly "remarkably sensitive to global forcings," and if "positive feedbacks predominate[d]," causing "the entire planet to be whipsawed between climatic states," as Hansen claims, especially in light of the tremendous ease with which he envisions hoards of earth's existing plant and animal species being driven to extinction by just a tad more warming than what the earth has already experienced?
(2) Hansen says "very little additional forcing is needed to cause ... an intensification of subtropical conditions that would greatly exacerbate water shortages in the American West and many other parts of the world, and likely render the semi-arid states from west and central Texas through Oklahoma, Kansas, Nebraska and the Dakotas increasingly drought prone and unsuitable for agriculture." With the significant enhancement of water use efficiency that is bestowed upon the planet's vegetation by rising atmospheric CO2 concentrations, however - which is a fact that has been demonstrated over and over in literally hundreds of laboratory and field experiments - it is by no means assured that Hansen's contention would be correct. In fact, it could well be false - and likely is, in our opinion - for the CO2-induced increase in plant water use efficiency that would accompany the rise in atmospheric CO2 concentration would allow the same amount of crop biomass and yield to be produced while transpiring a smaller quantity of water.
But don't take our word alone for this encouraging scenario, as Hansen expects you to take his word for the gloomy picture he paints; check out the pertinent scientific literature for yourself - much of which is summarized on our website (www.co2science.org) - as we do below for both a well-known C3 and C4 crop.
De Luis et al. (1999) grew alfalfa (a C3 plant) in controlled-environment chambers maintained at atmospheric CO2 concentrations of either 400 or 700 ppm for two weeks before imposing a two-week water treatment on them in which half of the plants were well-watered, with a soil moisture content approaching field capacity, and half of them were water-stressed, with a soil moisture content that was only 30% of field capacity. In spite of this significant difference in soil water status, they found that the water-stressed plants grown in air of 700 ppm CO2 produced 2.3 times more biomass than the well-watered plants grown in air of 400 ppm CO2, even though the water-stressed plants consumed less water than the plants growing in ambient air.
Likewise, utilizing a crop growth model tuned to local climate and crop characteristics, Grant et al. (2004) determined that a 50% increase in the air's CO2 content and an air temperature increase of 3°C would enhance the grain yield of sorghum (a C4 crop) by about 13%. Even more impressive was their finding that current high sorghum yields could be achieved with about "20% less irrigation water if these rises in temperature and CO2 were to occur." And because of the fact that the water use efficiencies of almost all plants rise significantly in response to increases in the air's CO2 content, almost all of the world's crops would behave similarly in this regard, producing equally as large yields as they do today - if not a little to a whole lot more - even under drier-than-current climate conditions in a CO2-enriched world of the future.
(3) Hansen says "there is increasing realization that sea level rise this century may be measured in meters [our italics] if we follow business-as-usual fossil fuel emissions." However, the real-world sea-level data we describe in the Sea Level Trends section of our critique indicate that sea level rise this century will more likely be measured in centimeters. And while Hansen says, with respect to sea level rise, that "the greatest threat is the likely demise of the West Antarctic ice sheet," the real-world measurements described in the Ice Sheet Disintegration section of our critique indicate that this particular source of sea level rise will likely be measured in millimeters.
(4) Hansen says "adaptation to a continually rising sea level is not possible," echoing the contentions of people throughout history who have emphatically claimed that this-or-that simply cannot be done. Time and again, however, almost all of them have been proven wrong, just as Hansen has already been proven wrong on this point by the many species of plants and animals that currently inhabit the earth, which have survived the many ups and downs of the global ocean when "the entire planet," as Hansen puts it, has been "whipsawed between climatic states." Truly, living organisms are incredibly adaptable, and man is the most adaptable of them all.
(5) Hansen says "the dangerous level of CO2 is at most 450 ppm, and it is probably less." It should be obvious to all, however, that this contention is pure speculation, especially since the key aspect of the claim resides in the highly subjective word "dangerous." This contention is totally incapable of being objectively adjudicated by science.
(6) Hansen says "doubling the amount of carbon dioxide in the atmosphere causes a global climate forcing similar in magnitude to that for a 2% increase of solar irradiance." All else being equal, this statement may not be far off the mark. However, it does not consider all of the negative biological feedbacks that the warming produced by the initial forcing might kick into motion, which could cause the long-term effective magnitude of the primary forcing to be significantly less than its initial value. Neither does it consider the cooling effects produced by increases in various biological processes that may be induced solely by the growth-enhancing effects of the increase in the air's CO2 content, which do not even require an impetus for warming in order to be put into play.
An example of the first of these ameliorative phenomena involves dimethylsulfide or DMS, which is derived from its algal precursor dimethylsulphoniopropionate. Very briefly, and rather simplistically, in response to an initial increase in temperature (caused by an increase in the air's CO2 content, for example), the climate-stabilizing mechanism begins with a warming-induced increase in the productivity of certain marine microalgae or phytoplankton, which leads to a greater production of oceanic DMS and its release to the atmosphere, which boosts the number of gas-to-particle conversions occurring there, increasing the atmosphere's population of cloud condensation nuclei and, ultimately, the albedos of marine stratus and altostratus clouds, via a narrowing of the cloud droplet spectrum and a decrease in the mean radius of the cloud droplets, both of which phenomena tend to counter the initial impetus for warming and thereby decrease the "all-else-being-equal" effect of the increase in the air's CO2 concentration, as originally suggested by Charlson et al. (1987).
Literally hundreds of peer-reviewed scientific papers have been published on this important subject over the past two decades, and recent work continues to demonstrate the great significance of this major negative feedback phenomenon. In one such study, Meskhidze and Nenes (2006) investigated the effects of ocean biological productivity on the microphysical and radiative properties of marine clouds over a large and seasonally-recurring phytoplankton bloom in the Southern Ocean in the vicinity of South Georgia Island, where the upwelling of nutrient-rich waters, as they describe it, "can support massive phytoplankton blooms, with chlorophyll-a concentrations more than an order of magnitude higher than the background."
In this ambitious endeavor, Meskhidze and Nenes used the Sea-viewing Wide Field-of-view Sensor to obtain the needed chlorophyll data and the Moderate Resolution Imaging Spectroradiometer to determine the effective radii of cloud condensation nuclei. These efforts revealed, in their words, that the "cloud droplet number concentration over the bloom was twice what it was away from the bloom, and cloud effective radius was reduced by 30%." In addition, they report that "the resulting change in the short-wave radiative flux at the top of the atmosphere was [a negative] 15 watts per square meter, comparable to the aerosol indirect effect over highly polluted regions," and, we might add, much greater locally than the opposite (positive) radiative forcing typically attributed to the combined increases in the concentrations of all greenhouse gases emitted to the atmosphere since the inception of the Industrial Revolution.
Other work in this area points to a similar anti-warming phenomenon that is produced by the effects of global change on the production of biogenic iodocarbons that are emitted by marine microalgae. O'Dowd et al. (2002), for example, report that emissions of iodocarbons from marine biota "can increase by up to five times as a result of changes in environmental conditions associated with global change," and that "increasing the source rate of condensable iodine vapors will result in an increase in marine aerosol and CCN [cloud condensation nuclei] concentrations of the order of 20-60%." Furthermore, they note that "changes in cloud albedo resulting from changes in CCN concentrations of this magnitude can lead to an increase in global raidative forcing similar in magnitude [our italics], but opposite in sign [our italics], to the forcing induced by greenhouse gases." Likewise, Smythe-Wright et al. (2006) report that "an increase in the production of iodocompounds and the subsequent production of CCNs would potentially result in a net cooling of the earth system and, hence, in a negative climate feedback mechanism, mitigating global warming."
In another recent experiment that dealt with both DMS and iodocarbons, Wingenter et al. (2007) studied the effects of atmospheric CO2 enrichment on marine microorganisms within 2-m-diameter polyethylene bags submerged to a depth of 10 meters in a fjord at the Large-Scale Facilities of the Biological Station of the University of Bergen in Espegrend, Norway, where three of the mesocosms were maintained at ambient CO2 levels (~375 ppm or base CO2), three at levels expected to prevail at the end of the current century (760 ppm or 2xCO2), and three at levels predicted for the middle of the next century (1150 ppm or 3xCO2). During the 25 days of the study, the researchers followed the development and subsequent decline of a bloom of the coccolithophorid Emiliania huxleyi, carefully measuring several physical, chemical and biological parameters. In doing so, they found that DMS production "followed the development and decline of the phytoplankton bloom," and that "DMS was 26% and 18% higher in the 2x and 3xCO2 mesocosms, respectively (days 0-17)," while chloroiodomethane had its peak concentration about 6-10 days after the chlorophyll-a maximum, and its abundance was 46% higher in the 2xCO2 mesocosms and 131% higher in the 3xCO2 mesocosms.
Based on the results of these several studies, the existence of the second type of biologically-induced marine cooling mechanism (which is CO2-driven as opposed to warming-driven) may readily be inferred from the significant increase in marine biological productivity that has been found to occur in a variety of atmospheric CO2 enrichment experiments (Riebesell et al., 1993; Hein and Sand-Jensen, 1997; Chen and Gao, 2004; Riebesell, 2004; Schippers et al., 2004; Lu et al., 2006; Levitan et al., 2007). In the case of the cyanobacterial diazotroph Trichodesmium growing in the laboratory in cultures in equilibrium with air of either high (900 ppm) or ambient (400 ppm) CO2 concentration, for example, Levitan et al. found that in the high CO2 treatment there was "a three- to four-fold increase in nitrogen fixation and a doubling of growth rates and biomass," while at the other end of the experimental spectrum - working "in the field" (i.e., at sea) - Riebesell (2004) reports that in the course of CO2 perturbation experiments conducted south of Bergen, Norway - where several 11-m3 enclosures moored to a floating raft were aerated with either normal (370-ppm) or CO2-enriched (710-ppm) air - a mixed phytoplankton community bloom developed in which "significantly higher net community production was observed under elevated CO2."
Consequently, in light of the facts that (1) increases in both water temperature and the atmosphere's CO2 concentration have been demonstrated to enhance marine biological productivity, that (2) increases in marine biological productivity have been demonstrated to lead to increases in the production of DMS and a number of iodocarbons, and that (3) increases in DMS and various iodocarbons have been demonstrated to be instrumental in the creation of (a) more, (b) brighter, and (c) longer-lasting clouds that ultimately lead to the reflection of more incoming solar radiation back to space, it is evident that a doubling of the atmosphere's CO2 concentration would not cause "a global climate forcing similar in magnitude to that for a 2% increase of solar irradiance," as Hansen claims it does.
(7) Hansen says that in lieu of using climate models, "a more definitive evaluation of climate sensitivity is provided by the earth's history," and in this he is correct. However, he goes on to claim that "empirical data for climate change over the past 700,000 years yield a climate sensitivity of 0.75°C for each W/m2 of forcing, or 3°C for a 4 W/m2 forcing," which just happens to be what most climate models imply for a doubling of the air's CO2 content. We, on the other hand, believe this result to be much too large, based on the empirical work of Idso (1998), who employed eight different empirical or "natural experiments" - covering time spans both shorter and longer than the one employed by Hansen and his colleagues - to derive a climate sensitivity that is almost an order of magnitude smaller than the sensitivity championed by Hansen.
(8) Hansen says "another ice age will never occur, unless humans go extinct." This is an incredibly forceful expression of essentially blind faith that is totally beyond the ability of the scientific enterprise to ever prove. It is another example of personal opinion, pure and simple, which is something that should be so indicated, especially within the context of scientific testimony regarding a scientific controversy.
(9) Hansen says "humans are now in control of global climate, for better or worse." This is yet another example of personal opinion masquerading as something that has been objectively determined by scientific methods.
(10) Hansen says "an ice age will never be allowed to occur if humans exist, because it can be prevented by even a 'thimbleful' of CFCs (chlorofluorocarbons), which are easily produced." Once again, Hansen exhibits blind faith in the capacity of humans to not only control global climate, but to do it easily. Many circumstances can be imagined where any number of linkages in the process leading to such an outcome could break down and thwart the stated outcome; and one could assert them just as forcefully as Hansen asserts his scenario (which contains no elucidation whatsoever).
(11) Hansen says "ignoring the climate problem at this time, for even another decade, would serve to lock in future catastrophic climatic change and impacts that will unfold during the remainder of this century and beyond." Once again, this claim is nothing more than personal opinion, which in our opinion is very likely false.
Consider the incredibly important fact (which we discuss in the Atmospheric Methane Concentrations section of our critique) that the historical increase in the atmosphere's methane concentration first slowed and then ceased to rise any further at the end of the 20th century. Then, consider the fact that the contribution of methane to anthropogenic climate forcing, to quote Dlugokencky et al. (2003), "is about half that from CO2 when direct and indirect components to its forcing are summed." Taken together, these two facts refute Hansen's claim; for the sudden disappearance of a climate forcing equivalent to 50% of that caused by the historic yearly increase in atmospheric CO2 concentration is vastly more than humanity could ever hope to bring to pass over the next decade and even far beyond.
But might not methane growth resume? We don't know ... and neither does Hansen! But if it were to resume, we believe it would likely not return to the rate of growth it exhibited previously. But it could! These are things we just don't know; and they demonstrate that one cannot make the kinds of emphatic declarations and calls-to-arms that Hansen makes throughout his far-from-dispassionate testimony.
In explaining our opinion of the matter - which is that significant further increases in the atmosphere's methane concentration are highly unlikely - we note that since the data presented by Khalil et al. (2007) show the mean global methane concentration to have been rising ever more slowly, in the mean, over the past quarter-century (to where it is now not rising at all), it seems to us to be rather unlikely that this well-established trend would suddenly experience the radical change that would be required to return it to the mean rate-of-rise it exhibited prior to its tapering off in the early 1980s. If anything, it seems more likely to us that the atmosphere's methane concentration might soon begin to decline, for the reason described in our discussion of Simpson et al.'s (2002) data in the Atmospheric Methane Concentrations section of our critique, as well as because of humanity's continuing efforts to prevent methane losses to the atmosphere at various places between its many points of extraction from the earth and its many points of usage.
(12) Hansen says "the planet is on the verge of dramatic climate change." Not so, we contend; for even if it had been "on the verge" - which is highly debatable - the recent stabilization of the atmosphere's methane concentration would have likely returned it from the precipice. And if the atmosphere's methane concentration actually begins to decline fairly soon - which scenario appears quite plausible - it would remove the planet even further from "the verge."
(13) Hansen says we "are forced [our italics] to find a way to limit atmospheric CO2 more stringently than has generally been assumed." This strident claim is pure opinion, and not just pure opinion, but bad opinion, in our opinion. Based on the disappearance of a yearly amount of climate forcing equivalent to half that produced by CO2, we can clearly be considerably less stringent in our attempts to limit CO2 emissions, for this reduction in anthropogenic forcing, which has occurred through no overt action on humanity's part, far exceeds anything we could ever hope to do for a very long time, as noted in our discussion of "Hansen says" item number 11.
(14) Hansen says "reduction of non-CO2 forcings provides some, but only moderate, flexibility in the CO2 ceiling." Considering what we report in the prior three "Hansen says" items, this claim is also seen to be erroneous, and for the same reasons.
(15) Hansen says "we cannot shrink from our moral responsibilities" in terms of what needs to be done "to preserve the planet for future generations," and in this declaration he is correct. Nevertheless, to be truly moral in this regard, and to truly "preserve the planet for future generations," we must act upon correct principles and in harmony with all of the pertinent facts; and after carefully analyzing Hansen's testimony, we are forced to conclude that his policy prescriptions are not consistent with these requirements.
Clearly, no one on either side of the debate over what to do - or not do - about rising atmospheric CO2 concentrations is any more or less "moral" than anyone else in this regard, unless that person promotes a view that he or she believes, or actually knows, to be inconsistent with what he or she perceives to be the truth; and if the latter is the case, that person is simply immoral. Consequently, although Hansen states the truth yet again when he says that if we "set in motion irreversible changes to the detriment of nature and humanity, it will be hard to explain our role to our children and grandchildren," it is equally true that if we implement the policies Hansen prescribes and they prove to be detrimental (which we believe they are), it will be equally "hard to explain our role to our children and grandchildren."
What we need, therefore, is much less talk about the motives of the various people and organizations who are providing input to international discussions about the causes and consequences of anthropogenic CO2 emissions and much more focusing of our collective energies on trying to discern the true nature of this most momentous of concerns. Hansen's testimony suggests that we already know enough to implement his policy prescriptions. We, on the other hand, believe that real-world evidence proves this view to be incorrect, and that appeals to morality by people on both sides of the issue will only make it more difficult to resolve remaining uncertainties. Clearly, only more - and more thorough - science will reveal the truth; and until the truth is known, morality cannot possibly indicate how we should act. Its invocation at this point in time, therefore, only tends to polarize the debate and reduce it to an emotional issue, when what is sorely needed is additional data discovery and truly objective rational analysis.
After a careful study of the claims made by James Hansen in his testimony of 26 April 2007 to the Select Committee of Energy Independence and Global Warming of the US House of Representatives, we find that much of what he contends is contradicted by real-world observations.
Although Hansen speaks of a sea level rise this century measured in meters, due to "the likely demise of the West Antarctic ice sheet," the most recent and comprehensive review of potential sea level rise due to contributions arising from the wastage of both the Antarctic and Greenland ice sheets suggests a century-long rise of only 35 millimeters, based on the results of 14 satellite-derived estimates of imbalances of the polar ice sheets that have been obtained since 1998. In addition, whereas Hansen claims that the rate of sea level rise is accelerating, century-scale data sets indicate that the mean rate-of-rise of the global ocean has either not accelerated at all over the latter part of the 20th century or has actually slowed.
Another of Hansen's claims that is at odds with reality is that atmospheric greenhouse gas concentrations are "skyrocketing," for several studies of methane (which has historically provided a climate forcing equivalent to approximately half that provided by CO2) have demonstrated that its atmospheric concentration actually stabilized several years ago and has ceased to rise further. This development - which was totally unanticipated by the Intergovernmental Panel on Climate Change at the time of its last major report, and which was vehemently denied to even be occurring when it was first observed - effectively repudiates Hansen's contentions about the need to act immediately to curtail anthropogenic CO2 emissions, for this unforeseen circumstance has already done more than humanity could ever hope to do in the foreseeable future in terms of reducing the atmosphere's radiative impetus for warming; and it has thereby given us considerable extra time to determine what the true status of earth's climate really is, as well as what we should, or should not, do about it.
So what is the "true status" of earth's climate? It is perhaps best understood by noting that the earth is not any warmer now - and is possibly a fair amount cooler - than it was at many other times in the past. These warmer-than-present periods include much of the Medieval Warm Period of a thousand years ago, most of the Climatic Optimum that held sway during the central portion of the current interglacial, and significant portions of all four of the prior interglacials, when - in all six cases - the air's CO2 concentration was much lower than it is today.
Why are these facts important? They are important because they demonstrate that today's temperatures are not in any way unusual, unnatural or unprecedented, contrary to what Hansen claims. In fact, today's temperatures are just what should be expected, as a result of the natural (non-CO2-induced) recovery of the planet from the global chill of the several-hundred-year-long Little Ice Age, which in many parts of the world was the coldest period of the current interglacial, and which was definitely not caused by a decline in atmospheric CO2 concentration in the centuries that preceded it, because there was no CO2 decline then, which further implies that a reversal of whatever did cause the Little Ice Age is likely what has led to its demise and the subsequent increase in mean global air temperature.
Hansen also foresees a warming-induced "extermination of a large fraction of plant and animal species," with many at high latitudes and altitudes being "pushed off the planet." However, as demonstrated by the scientific studies we cite, warming - especially when accompanied by an increase in the atmosphere's CO2 concentration - typically results in an expansion of the ranges of terrestrial plants and animals, leading to increases in biodiversity almost everywhere on the planet. Likewise, where Hansen sees nothing but "destruction of coral reefs and other ocean life" in response to a predicted CO2-induced acidification of the world's oceans, real-world observations suggest just the opposite.
One thing that Hansen does not foresee, however, is the shortfall in food production that may exist just a few short decades from now. Even considering hoped-for advancements in agricultural expertise and anticipated improvements in farming techniques, a number of knowledgeable scientists project there will be insufficient food to support the human population of the planet expected in the year 2050, unless humanity usurps all of the earth's remaining freshwater resources, as well as a good proportion of its cultivatable land, merely to grow what we will need to sustain ourselves at that point in time. These actions, if taken, will drastically reduce the amount of natural habitat available for the many plant and animal species with which we share the planet, insuring the extinctions of vast numbers of them, unless the air's CO2 content continues to rise, so that the crop productivity enhancement provided by CO2's aerial fertilization effect, along with the increase in crop water use efficiency provided by its anti-transpiration effect, can obviate the need for the land and water takings that will otherwise be required to meet the predicted shortfall in agricultural production.
In light of these several observations, it is clear there is another whole side to the CO2-climate issue in addition to the one described by Hansen; and the things that Hansen ignores totally alter the way the issue must be approached for a successful resolution of the multiple dilemmas confronting us. No longer can the actions that Hansen proposes be described as constituting a no-regrets insurance policy, as the world's climate alarmists typically characterize them. There are "regrets" associated with these policy proposals, and they may be even more horrific than the catastrophes imagined by Hansen. Consequently, it can be appreciated that the donning of the "cloak of morality" is not as easily accomplished in the real world of nature as it is in the virtual world of climate modeling, which continues to ignore many of the non-climatic effects of atmospheric CO2 enrichment to - we believe - the detriment of humanity and nature alike.
As a result of our analysis of Hansen's testimony, we find very little evidence to justify his policy prescriptions for dealing with what he calls a "dangerous climate change," but we find significant evidence for an impending world food supply-and-demand problem that may well prove even more devastating to the biosphere - including both humanity and "wild nature" - than what Hansen contends will occur in response to business-as-usual anthropogenic CO2 emissions. A complication introduced by this more recently recognized problem is that its solution would appear to involve our not allowing human-induced CO2 emissions to be restricted, which is just the opposite of what Hansen's policy prescriptions encourage. Clearly, the several interrelated aspects of this many-faceted conundrum demonstrate that it is vastly more complex than how Hansen has characterized it in his testimony and, therefore, that its solution is likely not to be found in the policies he prescribes.
Alley, R.B., Anandakrishnan, S., Dupont, T.K., Parizek, B.R. and Pollard, D. 2007. Effect of sedimentation on ice-sheet grounding-line stability. Science 315: 1838-1841.
Anandakrishnan, S., Catania, G.A., Alley, R.B. and Horgan, H.J. 2007. Discovery of till deposition at the grounding line of Whillans Ice Stream. Science 315: 1835-1838.
Anderson, J.B. 2007. Ice sheet stability and sea-level rise. Science 315: 1803-1804.
Bartlein, P.J., Webb, T., III. and Fleri, E. 1984. Holocene climatic change in the northern Midwest: Pollen-derived estimates. Quaternary Research 22: 361-374.
Bessat, F. and Buigues, D. 2001. Two centuries of variation in coral growth in a massive Porites colony from Moorea (French Polynesia): a response of ocean-atmosphere variability from south central Pacific. Palaeogeography, Palaeoclimatology, Palaeoecology 175: 381-392.
Bekki, S., Law, K.S. and Pyle, J.A. 1994. Effect of ozone depletion on atmospheric CH4 and CO concentrations. Nature 371: 595-597.
Bernabo, J.C. and Webb III, T. 1977. Changing patterns in the Holocene pollen record of northeastern North America: A mapped summary. Quaternary Research 8: 64-96.
Bischoff, W.W., Mackenzie, F.T. and Bishop, F.C. 1987. Stabilities of synthetic magnesian calcites in aqueous solution: Comparison with biogenic materials. Geochimica et Cosmochimica Acta 51: 1413-1423.
Brussaard, C.P.D., Gast, G.J., van Duyl, F.C. and Riegman, R. 1996. Impact of phytoplankton bloom magnitude on a pelagic microbial food web. Marine Ecology Progress Series 144: 211-221.
Buddemeier, R.W., Lkeypas, J.A. and Aronson, R.B. 2004. Coral Reefs & Global Climate Change: Potential Contributions of Climate Change to Stresses on Coral Reef Ecosystems. The Pew Center on Global Climate Change, Arlington, VA, USA.
Carricart-Ganivet, J.P. 2004. Sea surface temperature and the growth of the West Atlantic reef-building coral Montastraea annularis. Journal of Experimental Marine Biology and Ecology 302: 249-260.
Charlson, R.J., Lovelock, J.E., Andrea, M.O. and Warren, S.G. 1987. Oceanic phytoplankton, atmospheric sulfur, cloud albedo and climate. Nature 326: 655-661.
Chen, X. and Gao, K. 2004. Characterization of diurnal photosynthetic rhythms in the marine diatom Skeletonema costatum grown in synchronous culture under ambient and elevated CO2. Functional Plant Biology 31: 399-404.
Church, J.A., White, N.J., Coleman, R., Lambert, K. and Mitrovica, J.X. 2004. Estimates of the regional distribution of sea level rise over the 1950-2000 period. Journal of Climate 17: 2609-2625.
Clausen, C.D. and Roth, A.A. 1975. Effect of temperature and temperature adaptation on calcification rate in the hematypic Pocillopora damicornis. Marine Biology 33: 93-100.
Coles, S.L. and Coles. P.L. 1977. Effects of temperature on photosynthesis and respiration in hermatypic corals. Marine Biology 43: 209-216.
COHMAP Members. 1988. Climatic changes of the last 18,000 years: Observations and model simulations. Science 241: 1043-1052.
Cowling, S.A. 1999. Plants and temperature - CO2 uncoupling. Science 285: 1500-1501.
Crabbe, M.J.C., Wilson, M.E.J. and Smith, D.J. 2006. Quaternary corals from reefs in the Wakatobi Marine National Park, SE Sulawesi, Indonesia, show similar growth rates to modern corals from the same area. Journal of Quaternary Science 21: 803-809.
Davis, M.B., Spear, R.W. and Shane, L.C.K. 1980. Holocene climate of New England. Quaternary Research 14: 240-250.
De Luis, J., Irigoyen, J.J. and Sanchez-Diaz, M. 1999. Elevated CO2 enhances plant growth in droughted N2-fixing alfalfa without improving water stress. Physiologia Plantarum 107: 84-89.
Dickson, A.G. 1990. Thermodynamics of the dissociation of boric acid in synthetic seawater from 273.15 to 318.15K. Deep-Sea Research 37: 755-766.
Dickson, A.G. and Riley, J.P. 1979. The estimation of acid dissociation constants in seawater media from potentiometric titrations with strong base. I. The ionic product of water - KW. Marine Chemistry 7: 89-99.
Dlugokencky, E.J., Dutton, E.G., Novelli, P.C., Tans, P.P., Masarie, K.A., Lantz, K.O. and Madronich, S. 1996. Changes in CH4 and CO growth rates after the eruption of Mt. Pinatubo and their link with changes in tropical tropospheric UV flux. Geophysical Research Letters 23: 2761-2764.
Dlugokencky, E.J., Houweling, S., Bruhwiler, L., Masarie, K.A., Lang, P.M., Miller, J.B. and Tans, P.P. Atmospheric methane levels off: Temporary pause or a new steady-state? 2003. Geophysical Research Letters 30: 10.1029/2003GL018126.
Hansen, J.E. and Sato, M. 2001. Trends of measured climate forcing agents. Proceedings of the National Academy of Sciences USA 98: 14,778-14,783.
Dlugokencky, E.J., Masarie, K.A., Lang, P.M. and Tans, P.P. 1998. Continuing decline in the growth rate of the atmospheric methane burden. Nature 393: 447-450.
Dlugokencky, E.J., Walter, B.P., Masarie, K.A., Lang, P.M. and Kasischke, E.S. 2001. Measurements of an anomalous global methane increase during 1998. Geophysical Research Letters 28: 499-502.
Fine, M. and Tchernov, D. 2007. Scleractinian coral species survive and recover from decalcification. Science 315: 1811.
Francey, R.J., Manning, M.R., Allison, C.E., Coram, S.A., Etheridge, D.M., Langenfelds, R.L., Lowe, D.C. and Steele, L.P. 1999. A history of δ13C in atmospheric CH4 from the Cape Grim Air Archive and Antarctic firn air. Journal of Geophysical Research 104: 23,631-23,643.
Gnaiger, E., Gluth, G. and Weiser, W. 1978. pH fluctuations in an intertidal beach in Bermuda. Limnology and Oceanography 23: 851-857.
Grabherr, G., Gottfried, M. and Pauli, H. 1994. Climate effects on mountain plants. Nature 369: 448.
Grant, R.F., Kimball, B.A., Wall, G.W., Triggs, J.M., Brooks, T.J., Pinter Jr., P.J., Conley, M.M., Ottman, M.J., Lamorte, R.L., Leavitt, S.W., Thompson, T.L. and Matthias, A.D. 2004. Modeling elevated carbon dioxide effects on water relations, water use, and growth of irrigated sorghum. Agronomy Journal 96: 1693-1705.
Grigg, R.W. 1981. Coral reef development at high latitudes in Hawaii. In: Proceedings of the Fourth International Coral Reef Symposium, Manila, Vol. 1: 687-693.
Grigg, R.W. 1997. Paleoceanography of coral reefs in the Hawaiian-Emperor Chain - revisited. Coral Reefs 16: S33-S38.
Hansen, P.J. 2002. The effect of high pH on the growth and survival of marine phytoplankton: implications for species succession. Aquatic Microbiology and Ecology 28: 279-288.
Hansen, J., Sato, M., Ruedy, R., Lo, K., Lea, D.W. and Medina-Elizade, M. 2006. Global temperature change. Proceedings of the National Academy of Sciences USA 103: 14,288-14,293.
Hein, M. Sand-Jensen, K. 1997. CO2 increases oceanic primary production. Nature 388: 526-527.
Hofer, H.R. 1992. Veranderungen in der Vegetation von 14 Gipfeln des Berninagebietes zwischen 1905 und 1985. Ber. Geobot. Inst. Eidgenoss. Tech. Hochsch. Stift. Rubel Zur 58: 39-54.
Holgate, S.J. 2007. On the decadal rates of sea level change during the twentieth century. Geophysical Research Letters 34: 10.1029/2006GL028492.
Holgate, S.J. and Woodworth, P.L. 2004. Evidence for enhanced coastal sea level rise during the 1990s. Geophysical Research Letters 31: 10.1029/2004GL019626.
Hope, G.S., Peterson, J.A., Radok, U. and Allison, I. 1976. The Equatorial Glaciers of New Guinea. Balkema, Rotterdam.
Howat, I.M., Joughin, I. and Scambos, T.A. 2007. Rapid changes in ice discharge from Greenland outlet glaciers. Science 315: 1559-1561.
Huang, J., Pray, C. and Rozelle, S. 2002. Enhancing the crops to feed the poor. Nature 418: 678-684.
Huntley, B. and Prentice, C. 1988. July temperatures in Europe from pollen data 6000 years before present. Science 241: 687-690.
Idso, C.D. and Idso, K.E. 2000. Forecasting world food supplies: The impact of the rising atmospheric CO2 concentration. Technology 7S: 33-55.
Idso, K.E. and Idso, S.B. 1994. Plant responses to atmospheric CO2 enrichment in the face of environmental constraints: a review of the past 10 years' research. Agricultural and Forest Meteorology 69: 153-203.
Idso, S.B. 1998. CO2-induced global warming: a skeptic's view of potential climate change. Climate Research 10: 69-82.
Idso, S.B., Idso, C.D. and Idso, K.E. 2000. CO2, global warming and coral reefs: Prospects for the future. Technology 7S: 71-94.
Idso, S.B., Idso, C.D. and Idso, K.E. 2003. The Specter of Species Extinction: Will Global Warming Decimate Earth's Biosphere. Center for the Study of Carbon Dioxide... (http://www.co2science.org/../../../education/reports/extinction/extinction.php)
Idso, S.B. and Kimball, B.A. 2001. CO2 enrichment of sour orange trees: 13 years and counting. Environmental and Experimental Botany 46: 147-153.
Ishii, M., Kimoto, M. and Kachi, M. 2003. Historical ocean subsurface temperature analysis with error estimates. Monthly Weather Review 131: 51-73.
Jevrejeva, S., Grinsted, A., Moore, J.C. and Holgate, S. 2006. Nonlinear trends and multiyear cycles in sea level records. Journal of Geophysical Research 111: 10.1029/2005JC003229.
Jurik, T.W., Weber, J.A. and Gates, D.M. 1984. Short-term effects of CO2 on gas exchange of leaves of bigtooth aspen (Populus grandidentata) in the field. Plant Physiology 75: 1022-1026.
Kajiwara, K., Nagai, A. and Ueno, S. 1995. Examination of the effect of temperature, light intensity and zooxanthellae concentration on calcification and photosynthesis of scleractinian coral Acropora pulchra. J. School Mar. Sci. Technol. 40: 95-103.
Kearney, M.S. and Luckman, B.H. 1983. Holocene timberline fluctuations in Jasper National Park, Alberta. Science 221: 261-263.
Keller, F., Kienast, F. and Beniston, M. 2000. Evidence of response of vegetation to environmental change on high-elevation sites in the Swiss Alps. Regional Environmental Change 1: 70-77.
Khalil, M.A.K., Butenhoff, C.L. and Rasmussen, R.A. 2007. Atmospheric methane: Trends and cycles of sources and sinks. Environmental Science & Technology 10.1021/es061791t.
Klanderud, K. and Birks, H.J.B. 2003. Recent increases in species richness and shifts in altitudinal distributions of Norwegian mountain plants. Holocene 13: 1-6.
Kleypas, J.A., Buddemeier, R.W., Archer, D., Gattuso, J-P., Langdon, C., and Opdyke, B.N. 1999. Geochemical consequences of increased atmospheric carbon dioxide on coral reefs. Science 284: 118-120.
Korotky, A.M., Pletnev, S.P., Pushkar, V.S., Grebennikova, T.A., Raszhigaeva, N.T., Sahebgareeva, E.D. and Mohova, L.M. 1988. Development of Natural Environment of the Southern Soviet Far East (Late Pleistocene-Holocene). Nauka, Moscow, USSR.
Kullman, L. 2002. Rapid recent range-margin rise of tree and shrub species in the Swedish Scandes. Journal of Ecology 90: 68-77.
Kullman, L. 2007. Long-term geobotanical observations of climate change impacts in the Scandes of West-Central Sweden. Nordic Journal of Botany 24: 445-467.
Lacoul, P. and Freedman, B. 2006. Recent observation of a proliferation of Ranunculus trichophyllus Chaix. in high-altitude lakes of Mount Everest Region. Arctic, Antarctic and Alpine Research 38: 394-398.
Lassey, K.R., Lowe, D.C. and Manning, M.R. 2000. The trend in atmospheric methane δ13C and implications for constraints on the global methane budget. Global Biogeochemical Cycles 14: 41-49.
Levitan, O., Rosenberg, G., Setlik, I., Setlikova, E., Grigel, J., Klepetar, J., Prasil, O. and Berman-Frank, I. 2007. Elevated CO2 enhances nitrogen fixation and growth in the marine cyanobacterium Trichodesmium. Global Change Biology 13: 531-538.
Levitus, S., Stephens, C.M., Antonov, J.I. and Boyer, T.P. 2000. Yearly and Year-Season Upper Ocean Temperature Anomaly Fields, 1948-1998, U.S. Government Printing Office, Washington, DC.
Lindholm, T. and Nummelin, C. 1999. Red tide of the dinoflagellate Heterocapsa triquetra (Dinophyta) in a ferry-mixed coastal inlet. Hydrobiologia 393: 245-251.
Lombard, A., Cazenave, A., Le Traon, P.-Y. and Ishii, M. 2005. Contribution of thermal expansion to present-day sea-level change revisited. Global and Planetary Change 47: 1-16.
Lough, J.M. and Barnes, D.J. 1997. Several centuries of variation in skeletal extension, density and calcification in massive Porites colonies from the Great Barrier Reef: A proxy for seawater temperature and a background of variability against which to identify unnatural change. Journal of Experimental and Marine Biology and Ecology 211: 29-67.
Lough, J.B. and Barnes, D.J. 2000. Environmental controls on growth of the massive coral Porites. Journal of Experimental Marine Biology and Ecology 245: 225-243.
Lowe, D.C., Manning, M.R., Brailsford, G.W. and Bromley, A.M. 1997. The 1991-1992 atmospheric methane anomaly: Southern hemisphere 13C decrease and growth rate fluctuations. Geophysical Research Letters 24: 857-860.
Lu, Z., Jiao, N. and Zhang, H. 2006. Physiological changes in marine picocyanobacterial Synechococcus strains exposed to elevated CO2 partial pressure. Marine Biology Research 2: 424-430.
Lutaenko, K.A. 1993. Climatic optimum during the Holocene and the distribution of warm-water mollusks in the Sea of Japan. Palaeogeography, Palaeoclimatology, Palaeoecology 102: 273-281.
Macedo, M.F., Duarte, P., Mendes, P. and Ferreira, G. 2001. Annual variation of environmental variables, phytoplankton species composition and photosynthetic parameters in a coastal lagoon. Journal of Plankton Research 23: 719-732.
Mann, M., Amman, C., Bradley, R., Briffa, K., Jones, P., Osborn, T., Crowley, T., Hughes, M., Oppenheimer, M., Overpeck, J., Rutherford, S., Trenberth, K. and Wigley, T. 2003. On past temperatures and anomalous late-20th century warmth. EOS, Transactions, American Geophysical Union 84: 256-257.
Mann, M.E. and Jones, P.D. 2003. Global surface temperatures over the past two millennia. Geophysical Research Letters 30: 10.1029/2003GL017814.
McNeil, B.I., Matear, R.J. and Barnes, D.J. 2004. Coral reef calcification and climate change: The effect of ocean warming. Geophysical Research Letters 31: 10.1029/2004GL021541.
Medina, M., Collins, A.G., Takaoka, T.L., Kuehl, J.V. and Boore, J.L. 2006. Naked corals: Skeleton loss in Scleractinia. Proceedings of the National Academy of Science U.S.A. 103: 9096-9100.
Medina-Elizade, M. and Lea, D.W. 2005. The mid-Pleistocene transition in the tropical Pacific. Science 310: 1009-1012.
Meskhidze, N. and Nenes, A. 2006. Phytoplankton and cloudiness in the Southern Ocean. Science 314: 1419-1423.
Millero, F.J. 1995. Thermodynamics of the carbon dioxide system in the oceans. Geochimica et Cosmochimica Acta 59: 661-677.
Mucci, A. 1983. The solubility of calcite and aragonite in seawater at various salinities, temperatures, and one atmosphere total pressure. American Journal of Science 283: 780-799.
Nakicenovic, N., et al. 2000. IPCC Special Report on Emissions Scenarios. Cambridge University Press, Cambridge, UK.
Nie, B., Chen, T., Liang, M., Wang, Y., Zhong, J. and Zhu, Y. 1997. Relationship between coral growth rate and sea surface temperature in the northern part of South China Sea. Sci. China Ser. D 40: 173-182.
O'Dowd, C.D., Jimenez, J.L., Bahreini, R., Flagan, R.C., Seinfeld, J.H., Hameri, K., Pirjola, L., Kulmala, M., Jennings, S.G. and Hoffmann, T. 2002. Marine aerosol formation from biogenic iodine emissions. Nature 417: 632-636.
Overpeck, J.T. 1985. A pollen study of a late Quaternary peat bog, south-central Adirondack Mountains, New York. Geological Society of America Bulletin 96: 145-154.
Orr, J.C., Fabry, V.J., Aumont, O., Bopp, L., Doney, S.C., Feely, R.A., Gnanadesikan, A., Gruber, N., Ishida, A., Joos, F., Key, R.M., Lindsay, K., Maier-Reimer, E., Matear, R., Monfray, P., Mouchet, A., Najjar, R.G., Plattner, G.-K., Rodgers, K.B., Sabine, C.L., Sarmiento, J.L., Schlitzer, R., Slater, R.D., Totterdell, I.J., Weirig, M.-F., Yamanaka, Y. and Yool, A. 2005. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437: 681-686.
Pagani, M., Arthur, M.A. and Freeman, K.H. 1999. Miocene evolution of atmospheric carbon dioxide. Paleoceanography 14: 273-292.
Parmesan, C., Ryrholm, N., Stefanescu, C., Hill, J.K., Thomas, C.D., Descimon, H., Huntley, B., Kaila, L., Kullberg, J., Tammaru, T., Tennent, W.J., Thomas, J.A. and Warren, M. 1999. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 399: 579-583.
Parmesan, C. and Yohe, G. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421: 37-42.
Porter, S.C. and Orombelli, G. 1985. Glacial concentration during the middle Holocene in the western Italian Alps: Evidence and implications. Geology 13: 296-298.
Prather, M., et al. 2001. Atmospheric chemistry and greenhouse gases. In: Houghton, J.T., et al. (Eds.), Climate Change 2001: The Scientific Basis, Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK.
Raven, P.H. 2002. Science, sustainability, and the human prospect. Science 297: 954-959.
Reynaud, S., Ferrier-Pages, C., Boisson, F., Allemand, D. and Fairbanks, R.G. 2004. Effect of light and temperature on calcification and strontium uptake in the scleractinian coral Acropora verweyi. Marine Ecology Progress Series 279: 105-112.
Reynaud-Vaganay, S., Gattuso, J.P., Cuif, J.P., Jaubert, J. and Juillet-Leclerc, A. 1999. A novel culture technique for scleractinian corals: Application to investigate changes in skeletal δ18O as a function of temperature. Marine Ecology Progress Series 180: 121-130.
Riebesell, U. 2004. Effects of CO2 enrichment on marine phytoplankton. Journal of Oceanography 60: 719-729.
Riebesell, U., Wolf-Gladrow, D.A. and Smetacek, V. 1993. Carbon dioxide limitation of marine phytoplankton growth rates. Nature 361: 249-251.
Ritchie, J.C., Cwynar, L.C. and Spear, R.W. 1983. Evidence from north-west Canada for an early Holocene Milankovitch thermal maximum. Nature 305: 126-128.
Root, T.L., Price, J.T., Hall, K.R., Schneider, S.H., Rosenzweig, C. and Pounds, J.A. 2003. Fingerprints of global warming on wild animals and plants. Nature 421: 57-60.
Rubel, E. 1912. Pflanzengeographische Monographie des Berninagebietes. Engelmann, Leipzig, DE.
Santhanam, R., Srinivasan, A., Ramadhas, V. and Devaraj, M. 1994. Impact of Trichodesmium bloom on the plankton and productivity in the Tuticorin bay, southeast coast of India. Indian Journal of Marine Science 23: 27-30.
Saxe, H., Ellsworth, D.S. and Heath, J. 1998. Tree and forest functioning in an enriched CO2 atmosphere. New Phytologist 139: 395-436.
Schippers, P., Lurling, M. and Scheffer, M. 2004. Increase of atmospheric CO2 promotes phytoplankton productivity. Ecology Letters 7: 446-451.
Scoffin, T.P., Tudhope, A.W., Brown, B.E., Chansang, H. and Cheeney, R.F. 1992. Patterns and possible environmental controls of skeletogenesis of Porites lutea, South Thailand. Coral Reefs 11: 1-11.
Shepherd, A. and Wingham, D. 2007. Recent sea-level contributions of the Antarctic and Greenland Ice Sheets. Science 315: 1529-1532.
Simpson, I.J., Blake, D.R. and Rowland, F.S. 2002. Implications of the recent fluctuations in the growth rate of tropospheric methane. Geophysical Research Letters 29: 10.1029/2001GL014521.
Smythe-Wright, D., Boswell, S.M., Breithaupt, P., Davidson, R.D., Dimmer, C.H. and Eiras Diaz, L.B. 2006. Methyl iodide production in the ocean: Implications for climate change. Global Biogeochemical Cycles 20: 10.1029/2005GB002642.
Stanley Jr., G.D. 2003. The evolution of modern corals and their early history. Earth Science Reviews 60: 195-225.
Stanley Jr., G.D. and Fautin, D.G. 2001. The origins of modern corals. Science 291: 1913-1914.
Taira, K. 1975. Temperature variation of the "Kuroshio" and crustal movements in eastern and southeastern Asia 7000 years B.P. Palaeogeography, Palaeoclimatology, Palaeoecology 17: 333-338.
Tilman, D., Cassman, K.G., Matson, P.A., Naylor, R. and Polasky, S. 2002. Agricultural sustainability and intensive production practices. Nature 418: 671-677.
Tilman, D., Fargione, J., Wolff, B., D'Antonio, C., Dobson, A., Howarth, R., Schindler, D., Schlesinger, W.H., Simberloff, D. and Swackhamer, D. 2001. Forecasting agriculturally driven global environmental change. Science 292: 281-284.
Velitchko, A.A. and Klimanov, V.A. 1990. Climatic zonality of the northern hemisphere 5 or 6 thousand years B.P. Proceedings of the USSR Academy of Sciences, Geographical Series, 5: 38-52.
Virtanen, R., Eskelinen, A. and Gaare, E. 2003. Long-term changes in alpine plant communities in Norway and Finland. In: Nagy, L., Grabherr, G., Korner, C. and Thompson, D.B.A. (Eds.), Alpine Biodiversity in Europe. Springer, Berlin, Germany, pp. 411-422.
Waggoner, P.E. 1995. How much land can ten billion people spare for nature? Does technology make a difference? Technology in Society 17: 17-34.
Wallace, J.S. 2000. Increasing agricultural water use efficiency to meet future food production. Agriculture, Ecosystems & Environment 82: 105-119.
Walther, G.-R., Beissner, S. and Burga, C.A. 2005. Trends in the upward shift of alpine plants. Journal of Vegetation Science 16: 541-548.
Webb, T. 1985. Holocene palynology and climate. In: Paleoclimate Analysis and Modeling. A.D. Hecht (Ed.). Wiley-Interscience, New York, NY, pp. 163-196.
Webb, T., Bartlein, P.J. and Kutzbach, J.E. 1987. Climatic change in eastern North America during the past 18,000 years: Comparisons of pollen data with model results. In: North America and Adjacent Oceans During the Last Deglaciation. W.F. Ruddiman and H.E. Wright, Jr. (Eds.), The Geology of North America, v. K-3. Geological Society of America, Boulder, CO, pp. 447-462.
Wijmstra, T.A. 1978. Paleobotany and climatic change. In: Climatic Change. J. Gribbin (Ed.), Cambridge University Press, New York, NY.
Wingenter, O.W., Haase, K.B., Zeigler, M., Blake, D.R., Rowland, F.S., Sive, B.C., Paulino, A., Thyrhaug, R., Larsen A., Schulz, K., Meyerhofer, M. and Riebesell, U. 2007. Unexpected consequences of increasing CO2 and ocean acidity on marine production of DMS and CH2CII: Potential climate impacts. Geophysical Research Letters 34: 10.1029/2006GL028139.